지질다당류가 비강상피세포의 Aspergillus fumigatus 생물막 형성에 미치는 영향
Effect of Lipopolysaccharide on Development of Aspergillus fumigatus Biofilm on Nasal Epithelial Cells
Article information

Trans Abstract
Background and Objectives
Aspergillus fumigatus is one of the common causes of fungal airway inflammatory diseases and lipopolysaccharide (LPS) acts as key regulator of airway inflammation. In addition, bacterial and fungal biofilm commonly coexist in chronic rhinosinusitis. In this study, we evaluated the effect of LPS on the development of A. fumigatus biofilm formation on the nasal epithelial cells.
Materials and Method
Primary nasal epithelial cells were cultured with A. fumigatus conidia with or without LPS for 5 days. The production of interleukin (IL)-6, IL-8, and transforming growth factor (TGF)-β1 from nasal epithelial cells was determined by enzyme-linked immunosorbent assay. The effects of LPS on A. fumigatus biofilm formation were determined using biofilm dry weight, and crystal violet, concanavalin A, safranin staining, and confocal scanning laser microscopy.
Results
LPS and A. fumigatus significantly enhanced the production of I L-6, I L-8, and TGF-β1 from nasal epithelial cells. A. fumigatus can form biofilm on primary nasal epithelial cells, and this significantly increased in a time-dependent manner when cocultured with LPS, the dry weight, concanavalin A, and safranin staining.
Conclusion
The exposure of A. fumigatus to LPS enhanced the formation of biofilms. The coexistence of LPS and A. fumigatus enhanced fungal biofilm formation and this may be associated with the development of recalcitrant airway inflammatory diseases.
서 론
만성 부비동염은 바이러스와 세균 감염이 주된 원인인 급성 부비동염과 달리 알레르기, 점액섬모수송 이상, 면역장애, 해부학적 요인 등 다양한 원인을 가지는 다인성 질환이다[1]. 분자생물학의 발전으로 1990년대에 들어 만성 부비동염의 발병에 진균의 역할에 대한 관심을 가지게 되었다[2]. 진균들이 비강상피세포, 섬유아세포, 호산구 등의 면역세포와 상호작용을 보인다는 연구들이 지속적으로 보고되고 있다[3,4]. 진균이 호흡기 점막의 선천성 혹은 후천성 면역반응을 유발하여 다양한 염증성 질환의 발병에 관여하며, 만성 부비동염의 발병과 연관하여서도 다양한 연구들이 진행되고 있다. 최근에는 호흡기 점막 진균총(mycobiome)의 역할에 대한 연구 또한 활발히 진행되고 있으며, 진균총의 변화 또한 호흡기 질환을 유발할 수 있음이 보고되고 있다[5,6].
생물막은 미생물에서 만들어진 세포외 고분자 물질(extracellular polymeric substances)과 세포외 기질(extracellular matrix)을 바탕으로 하는 미생물 공동체로 숙주의 방어 기전을 회피하고 약물에 대한 저항성을 가지게 하여 치료에 반응하지 않는 불응성 감염에 관여한다. 생물막은 세균뿐 아니라 진균에 의해서도 형성된다. 만성 부비동염의 경우 44%-92%에서 세균성 생물막과 진균성 생물막이 함께 존재하고 있으며, 진균성 생물막은 호산구성 점액에서 흔히 발견된다[7]. 호흡을 통해 기도로 들어온 진균 포자들이 기도 점막의 선천성 방어기전을 통해 제거되지 못하는 경우 점막 내에서 발아하여 균사체를 형성하게 되고 세포외 기질과 다당류와의 상호작용을 통해 진균성 생물막을 형성하게 된다. 진균은 호흡 점막의 방어기전이 정상적으로 작동하는 경우 생물막을 형성하기 어려우나 다양한 원인, 즉 호흡기 점막의 물리적 손상이나 세균성 혹은 항원에 의한 점막의 손상이 있는 경우 진균성 생물막이 형성될 수 있다. Staphylococcus aureus와 진균을 함께 비강 점막에 처리하는 경우 진균성 생물막이 형성될 수 있음이 보고되었다[8]. 대표적 진균성 만성부비동염인 진균구의 경우 세균성 감염을 동반하는 경우가 많으며, 세균성 생물막과 진균성 생물막이 공존하는 것을 흔히 발견할 수 있다[9].
지질다당류(lipopolysaccharide, LPS)는 그람음성균의 외막을 형성하는 세균성 내독소로 호흡기 염증성 질환 발병에 관여한다. 그람음성균은 약물이나 수술적 치료에 반응하지 않는 불응성 만성부비동염의 원인으로 알려져 있으며, LPS만으로도 만성부비동염을 유발할 수 있다[10]. Aspergillus fumigatus는 호흡기 진균성 질환과 부비동 진균구 등의 주된 원인으로 비강상피세포와 함께 배양하는 경우 진균성 생물막이 형성될 수 있다[11]. 본 연구는 LPS와 A. fumigatus 포자의 합동배양이 진균성 생물막의 형성에 영향을 미칠 수 있는지 확인해 보고자 진행하였다.
재료 및 방법
A. fumigatus 포자 수집
A. fumigatus는 American Type Culture Collection (Rockville, MD, USA)에서 구입하였다. Potato dextrose corn meal agar 배지에서 배양 후 진균 포자를 다음과 같이 분리하였다. 배양된 진균을 0.05% tween 20을 첨가한 5 mL의 인산완충식염수(phosphate buffer solution, PBS)로 세척하였으며, 얻어진 세척액을 1000 rpm으로 5분간 원심분리 후 40 μm의 여과기에 통과시켜 포자를 획득하였다. 2×107/mL의 포자를 45°C에서 건조시킨 후 -80°C에서 보관하였다.
비강상피세포의 분리 배양과 A. fumigatus 처리
비강상피세포는 비중격 만곡증 환자의 하비갑개에서 분리하여 사용하였다(n=10, 남자 6명, 여자 4명, 평균 나이 46.2±19.2). 대상자의 선택은 알레르기비염이 동반되지 않고, 수술 전 4주 이상 항생제, 항히스타민제, 국소 및 경구용 스테로이드제재 등의 약물 치료를 시행하지 않은 예로 하였으며, 대상자들은 저자 소속 병원의 임상윤리위원회 승인하에 진행되었다(CR-20-200-L). 비점막은 penicillin 100 UI/mL, streptomycin (SM) 100 μg/mL, amphotericin-B 2 μg/mL을 포함한 Ham’s F-12 배양액(GIBCO, Grand Island, NY, USA)으로 세척 후 dispase (GIBCO)로 처리하였으며, 37°C에서 trypsin/ethylenediamine tetraacetic acid로 10분간 배양하여 상피세포를 박리하였다. 박리된 세포들은 10 cm 접시에 1시간 두어 섬유모세포, 근육세포, 혈관 내피세포를 제거하였으며, 이렇게 얻어진 상피세포는 Pc, SM, Amp-B, glutamine 150 μg/mL, transferrin 5 μg/mL, insulin 5 UI/mL, epithelial growth factor 25 ng/mL, endothelial cell growth supplement 15 μg/mL, triiodothyronin 200 pM, hydrocortisone 100 nM, fetal calf serum 15%을 포함한 Ham’s F-12에 넣고, 37°C, 5% CO2에서 배양하였다.
A. fumigatus와 LPS에 의한 비강상피세포의 생존능력 변화 측정
A. fumigatus와 LPS가 비강상피세포 분화에 미치는 영향은 CellTiter-96Ⓡ aqueous cell proliferation assay kit (Promega, Madison, WI, USA)를 사용하여 측정하였다. 비강상피세포와 함께 다양한 농도의 A. fumigatus (1×102-1×104/mL)와 LPS (1-100 μg/mL)로 37°C, 5% CO2 상태에서 72시간 처리하였다. 이후 테트라졸륨 화합물과 오웬 시약을 첨가하여 세포 산화 환원효소 작용으로 세포 내 미트콘드리아에 의해 불용성 포르마잔으로 환원시킨다. 포르마잔은 생존 세포의 수를 반영해 준다. 이후 490 nm 파장의 형광 판독기를 사용하여 흡광도를 평가한다.
비강상피세포에서 만들어지는 interleukin (IL)-6, IL-8와 transforming growth factor (TGF)-β1 측정
A. fumigatus 처리 후 24, 48, 72시간에 세포배양 상등액을 획득하여 -70°C에 보관하였다. IL-6, IL-8, TGF-β1의 농도는 효소 결합 면역 협착 분석 키트를 이용하여 측정하였다(R&D system, Minneapolis, MN, USA). IL-6와 IL-8의 측정 민감도는 2 pg/mL였고 TGF-β1은 15 pg/mL였다.
A. fumigatus 생물막 확인
비강상피세포가 80%-90%의 confluence가 형성되면 A. fumigatus (1×102/mL)로 2시간 처리 후 10 ug/mL의 LPS로 5일간 배양하였다. 비강상피세포와 함께 배양된 A. fumigatus 생물막의 존재와 LPS의 영향은 다음의 방법으로 확인하였다. 1) 건조한 생물막의 무게는 배양 접시의 진균을 긁어 채취한 후 0.45 μm 크기의 셀룰로스 질산염 필터(Sartorius, Go¨ttingen, Germany)로 여과 후 40°C에서 건조 후 무게를 측정하였다. 2) Mowat 방법을 이용한 진균 생물막의 정량적 분석을 시행하였다[12]. A. fumigatus 배양 접시에 0.5% crystalviolet 액을 넣고 5분 처리 후 PBS로 세척하였으며, 100 μL의 95% 에탄올을 넣고 1분간 처리하였다. 생물막의 정량 분석을 위해 570 nm 파장을 이용하여 분광 광도계(FLUOstar OPTIMA; BMG LABTECH, Oldenburg, Germany)로 측정하였다. 3) 진균 생물막의 세포외 기질의 확인을 위해 A. fumigatus 배양 접시에 50 μL의 safranin 용액을 넣고 72시간 동안 처리하여 진균 생물막을 형성하는 다당류를 광학 밀도 492 nm의 분광 광도계로 측정하였다. 4) 진균 생물막을 형성하는 세포외 기질의 정량적 분석은 100 μL의 succinylated concanavalin A와 Alexa Fluor 488 (Invitrogen, Carlsbad, CA, USA)로 37°C에서 45분간 처리 후 485 nm의 여기 파장과 520 nm의 방출 파장에서 형광 강도를 측정하였다. 5) 비강상피세포와 A. fumigatus의 합동 배양 후 생물막을 주사형공초점 레이저 현미경(confocal scanning laser microscopy)을 이용하여 확인하였다. 100 μM의 FUN-1 (Invitrogen)과 100 μg/mL의 concanavalin A를 넣고 암실에서 45분간 처리하였다. PBS로 세척 후 NikonA1 공초점 현미경(Kikon, Tokyo, Japan)을 이용하여 영상을 분석하였다. 붉은 색은 대사가 활발히 이루어지고 있는 세포이며, 녹색은 세포벽을 형성하는 다당류, 노란색은 두 가지 조건을 동시에 충족하는 경우 나타나는 형광색이다.
통계 분석
각각의 실험은 5회 이상 반복하여 그 결과를 얻어 평균±표준오차로 표현하였으며, 두 군간의 비교 분석은 Student t-test를 사용하였고, 두 군 이상의 비교는 one-way analysis를 시행 후 Tukey’s test로 사후 검정하였다(SPSS ver. 25.0; IBM Corp., Armonk, NY, USA). 통계학적 유의수준은 p<0.05로 하였다.
결 과
A. fumigatus와 LPS의 비강상피세포 생존 능력에 미치는 영향
비강상피세포를 LPS 1 μg/mL, 10 μg/mL, 100 μg/mL로 72시간 동안 처리하여 세포 생존을 확인하였다. LPS 1 μg/mL과 10 μg/mL의 경우 배양 72시간까지 상피세포의 변화가 없었으나, LPS 100 μg/mL의 경우 배양 24시간 이후부터 세포 생존 능력이 의미있게 감소하였다(Fig. 1A). 이후 연구에서는 LPS 10 μg/mL을 이용하여 연구를 진행하였다.
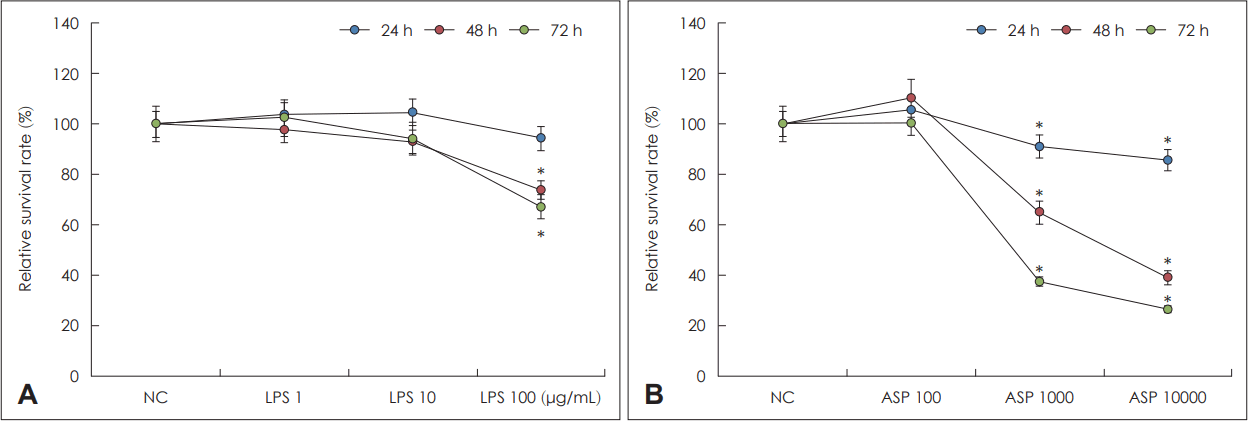
Cell viability effect of lipopolysaccharide (LPS) and Aspergillus fumigatus (ASP) on primary nasal epithelial cells. A: Relative survival of nasal epithelial cells, treated with 0 to 100 μg/mL of LPS for 72 h. B: Relative survival of nasal epithelial cells, treated with 0 to 1×104/mL of ASP conidia. Results are expressed as mean±standard error. *p<0.05 compared to negative control (NC), n=5.
A. fumigatus는 1×102/mL, 1×103/mL, 1×104/mL 포자를 비강상피세포와 함께 배양한 경우 1×102/mL의 경우 배양 72시간까지 상피세포의 생존 능력에 변화가 없었으나, 1×103/mL, 1×104/mL의 경우 배양 24시간부터 생존 능력이 의 미있게 감소됨이 확인되어, 이후 연구에서는 A. fumigatus 1×102/mL로 연구를 진행하였다(Fig. 1B).
A. fumigatus와 LPS의 비강상피세포의 IL-6, IL-8, TGF-β1 생성에 미치는 영향
비강상피세포에서 만들어지는 IL-6, IL-8, TGF-β1의 정량적 분석을 enzyme-linked immunosorbent assay법으로 측정하였다. 배양 24시간 경과한 경우 A. fumigatus, LPS 혹은 이 두 가지를 함께 처리한 경우 IL-6, IL-8, TGF-β1의 생성이 대조군에 비해 유의하게 증가하였다. 배양 48시간이 경과한 경우 LPS, A. fumigatus와 LPS를 함께 처리한 경우 세 가지 사이토카인의 생성이 유의하게 증가하였으며, 배양 72시간의 경우 IL-6는 LPS로 처리한 경우, IL-8은 A. fumigatus, A. fumigatus와 LPS를 함께 처리한 경우, TGF-β1은 A. fumigatus와 LPS를 함께 처리한 경우 대조군에 비해 유의하게 증가하였다(Fig. 2).
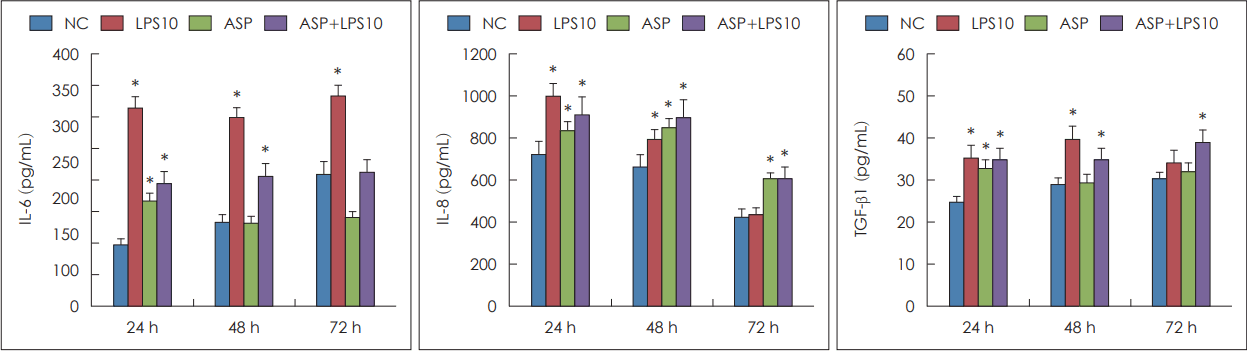
Effect pf lipopolysaccharide (LPS) and Aspergillus fumigatus (ASP) conidia on chemical mediator production in primary nasal epithelial cells. 10 μg/mL LPS, 1×102/mL ASP, or LPS and ASP enhanced interleukin (IL)-6, IL-8, and transforming growth factor (TGF)-β1 production. Results are expressed as mean±standard error. *p<0.05 compared to negative control (NC), n=7.
LPS가 비강상피세포와 함께 배양된 A. fumigatus 생물막 형성에 미치는 영향
A. fumigatus와 LPS를 비강상피세포와 함께 배양한 경우 생물막의 무게는 배양 1일째(3.0±0.7 mg)부터 비강상피세포 없이 A. fumigatus 단독 배양(1.3±0.4 mg)하거나, 비강상피세포와 A. fumigatus 함께 배양한 경우(1.6±0.4 mg)보다 유의하게 무거웠으며, 생물막의 무게는 배양시간의 경과에 따라 지속적으로 증가하였다(Fig. 3A). 생물막의 세포외 기질의 존재와 생존능력을 확인하기 위해 시행한 crystal violet 염색은 비강상피세포와 A. fumigatus를 함께 배양한 경우 유의하게 발현이 높았으며, LPS를 추가하는 경우 더욱 강하게 발현됨을 확인할 수 있었다(Fig. 3B). 진균 세포벽에 염색되는 concanavalin A의 경우 A. fumigatus와 LPS를 비강상피세포와 함께 배양한 경우 배양 1일째(4.3±1.4)부터 비강상피세포 없이 A. fumigatus 단독 배양(0.6±0.3)하거나, 비강상피세포와 A. fumigatus 함께 배양한 경우(0.8±0.4)보다 강하게 염색되었으나, LPS없이 배양하는 경우의 두 군간에는 차이를 보이지 않았다(Fig. 3C). 생물막 존재를 간접적으로 나타낼 수 있는 safranin 염색의 경우 A. fumigatus와 LPS를 비강상피세포와 함께 배양한 경우 배양 1일째(6.1±1.8)부터 다른 두 군보다 강하게 염색되었으며, 비강상피세포와 A. fumigatus를 함께 배양한 경우(1.3±0.5)에도 배양 1일째부터 A. fumigatus 단독의 배양의 경우(0.4±0.3)보다 강하게 염색되었다(Fig. 3D).
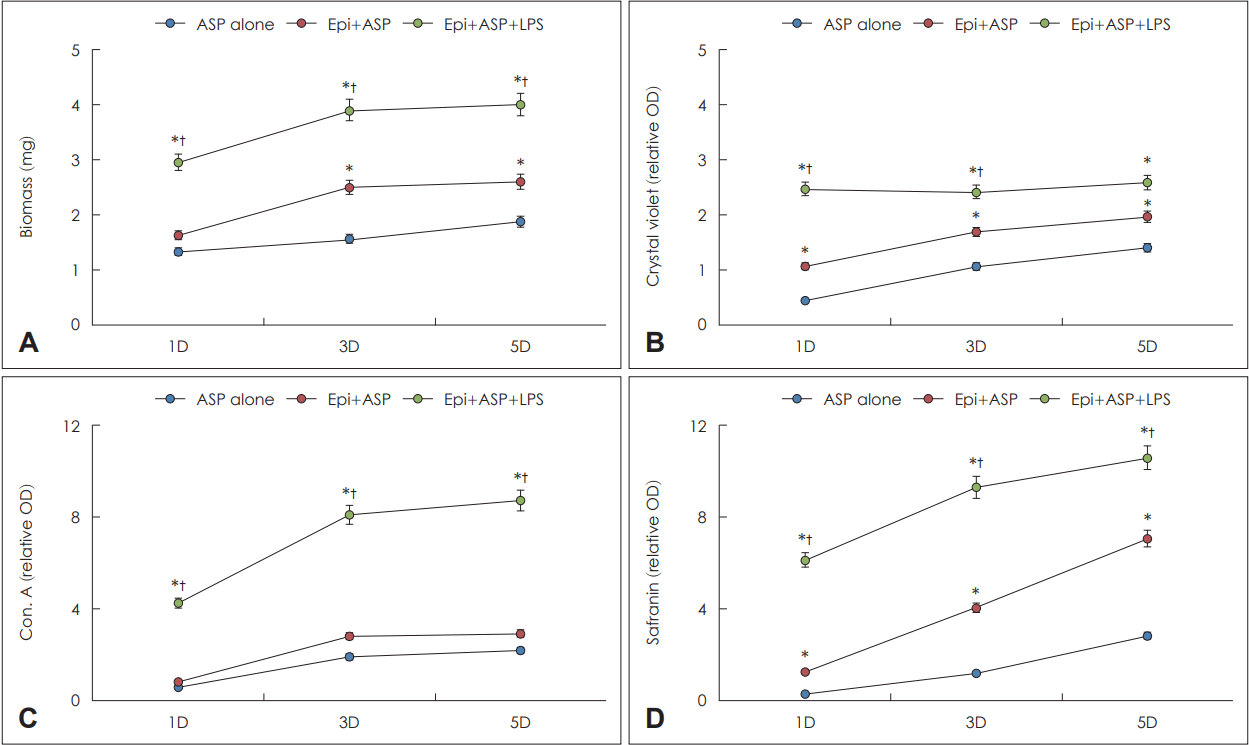
Quantification of Aspergillus fumigatus (ASP) biofilm in coculture of lipopolysaccharide (LPS) in primary nasal epithelial cells (Epi). Biofilm dry weight, concanavalin A (Con. A) and safranin staining intensity significantly increased in a time-dependent manner (A, C, D). Also, coculture of ASP and LPS in Epi (Epi+ASP+LPS) significantly increased the biofilm dry weight, Con. A, and safranin intensity compared with ASP cultured with Epi (Epi+ASP) or without Epi (ASP alone) (A, C, D). Crystal violet intensity also significantly increased in Epi+ASP+LPS compared with ASP alone or Epi+ASP (B). Results are expressed as mean ¡¾ standard error. *p<0.05 compared with ASP alone; †p<0.05 compared with Epi+ASP, n=5. D, day; OD, optical density.
A. fumigatus 생물막의 주사형 공초점 레이저 현미경을 이용한 확인
진균 생물막을 확인하기 위해 진균 세포벽에 존재하는 탄수화물과 반응하는 concanavalin A와 대사가 활발하게 이루어지는 세포질에 염색되는 FUN-1 염색을 시행하였다. Concanavalin A의 경우 A. fumigatus와 LPS를 비강상피세포와 함께 배양한 경우 배양 1일째(382.7±136.1)는 다른 두 군보다 강하게 염색되었으며(59.8±18.0과 103.1±34.1), 배양 3일째에는 비강상피세포와 A. fumigatus를 함께 배양한 경우 LPS가 생물막 형성에 추가적인 영향을 미치지 못하였으며(Fig. 4A-C), 배양 5일째는 세 군간의 차이를 보이지 않았다. Fun-1 염색의 경우 배양 1일째에는 세 군간의 차이를 보이지 않았으나, 3일째부터 A. fumigatus와 LPS를 비강상피 세포와 함께 배양한 경우 다른 두 군보다 강하게 염색되었으며, 비강상피세포와 A. fumigatus를 함께 배양한 경우에도 배양 3일째부터 A. fumigatus 단독의 배양의 경우보다 강하게 염색되었다(Fig. 4D and E).
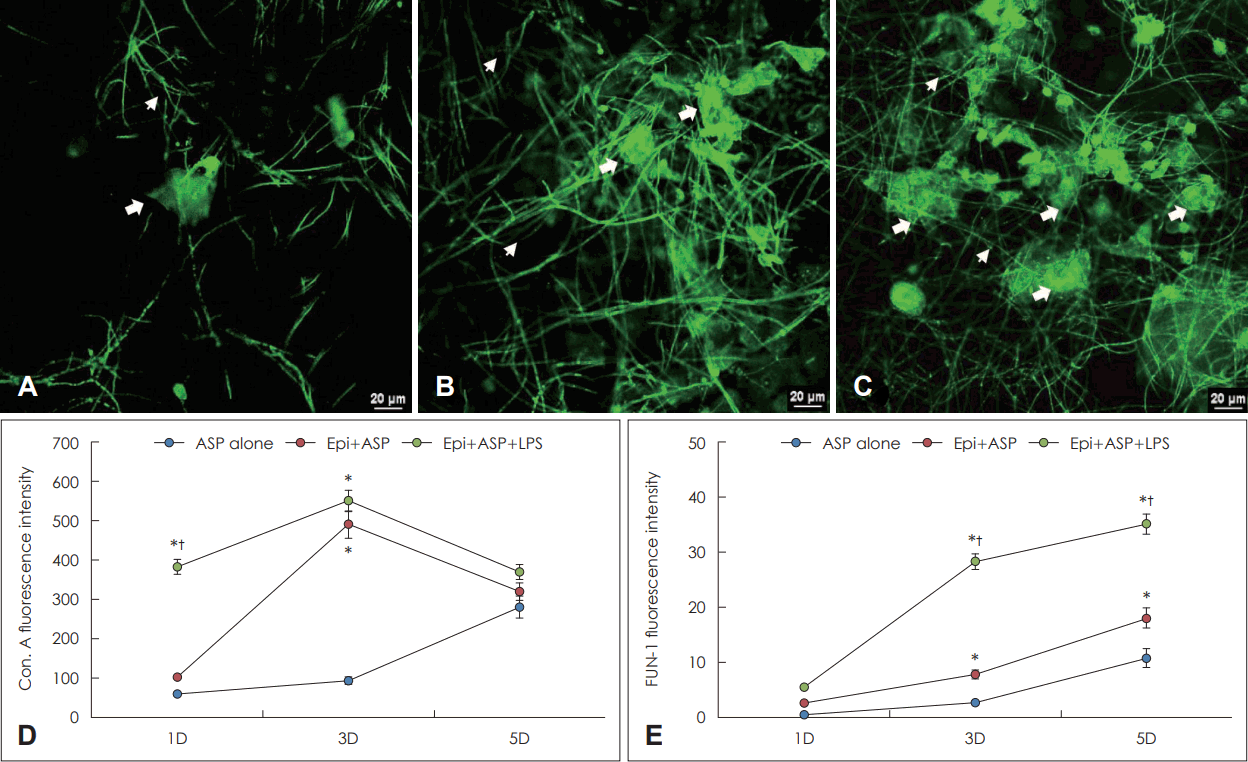
Confocal scanning laser microscopy findings of Aspergillus fumigatus (ASP) treated with lipopolysaccharide (LPS). A-C: Representative inverted microscopic and confocal scanning laser microscopic findings at day 3 (3D) coculturing of ASP alone (A), ASP on epithelial cells (B), and ASP with LPS on nasal epithelial cells (C). The arrow indicates concanavalin A (Con. A) stain binding to the fungal cell wall polysaccharide and extracellular matrix, and the arrowhead indicates FUN-1 stain binding to metabolically active fungi. When ASP was incubated with primary nasal epithelial cells, the fluorescent intensity of Con. A (D) and FUN-1 (E) increased. The coculture of ASP and LPS significantly enhanced the Con. A and FUN-1 intensity at 3D. Results are expressed as mean±standard error. *p< 0.05 compared with ASP alone; †p<0.05 compared with Epi+ASP, n=5.
고 찰
Aspergillus는 만성부비동염의 비즙에서 배양되며 진균구의 주된 원인균으로 진균성 비부비동 질환에서 흔히 발견되며, 아스페르길루스증(aspergillosis)이나 기관지 천식 환자의 기도에서도 흔히 발견된다[13-15]. 호흡을 통해 하루 수백 개의 A. fumigatus 포자가 흡입되어 호흡기 점막과 접촉하게 되며, 선천성 혹은 후천성 면역반응을 통해 제거되거나 다양한 염증반응을 유발하게 된다. A. fumigatus는 호흡상피세포의 dectin-1 수용체 결합하여 MyD88/NF-κB 혹은 PI3K/p38/ERK1/2 신호전달체계의 연쇄반응을 통해 각종 사이토카인과 화학매개물질의 생산을 유발한다[16]. 비강상피세포의 protease activated receptor, toll-like receptor나 C-type lectin-like receptor와 같은 패턴 인식 수용체와 결합하여 NF-κB와 MAPK 등의 신호전달체계를 통해 상피세포의 활성화를 유도하여 국소 염증반응을 유지 악화시킬 수 있다[4,17]. 염증성 사이토카인인 IL-6, 호중구 화학주성인자인 IL-8, 호흡상피세포의 세포외 기질 형성과 상피 간엽이행을 유도하는 TGF-β1은 A. fumigatus 포자 처리 24시간부터 모두 증가하였으며, IL-8의 경우 배양 72시간까지 지속적으로 증가하였다. 그 원인을 정확히 설명하기 어려우나 IL-8의 증가는 외부에서 침입한 병원성 물질의 제거에 관여하는 호중구의 이주를 유도하여 진균에 대한 인체 방어기전을 유지, 강화시킬 수 있을 것이다. 그람음성균 세포벽의 주성분인 LPS 또한 비강점막의 화학매개물질과 점액유전자의 발현을 증가시키는 것으로 알려져 있으며, 본 연구에서도 유사하게 IL-6, IL-8, TGF-β의 생성을 유의하게 증가시켰다[18]. 하지만 상피세포를 진균 포자와 LPS를 함께 처리한 경우 사이토카인 생성의 상승효과를 보이지 않았는데 이는 진균 포자와 LPS 함께 처리하는 경우 진균 생물막의 형성에 의한 대사활동의 감소에 기인하거나, 두 가지 물질이 동시에 상피세포 작용하여 세포독성을 증가시킴에 기인할 수 있을 것이다.
호흡기 감염에 있어 세균성 생물막이 항생제 내성 혹은 치료에 잘 반응하지 않는 염증성 질환의 원인으로 알려지면서 진균성 생물막에 대해서도 관심을 가지게 되었다. 진균성 생물막의 경우도 비강 세척과 같은 물리적 치료나 항진균제로 치료가 되지 않는 불응성 질환의 원인으로 임상적 의의를 가지고 있다[19]. A. fumigatus 생물막은 예각을 이루는 균사의 평형 혹은 교차 형태의 군집이 세포외 기질과 세포외 고분자 물질로 구성되어 부유 형태의 진균보다 대사활동이 감소하여 항진균제에 대한 강한 저항성을 가지게 된다[12]. A. fumigatus 포자의 부착과 발아가 이루어지면서 세포외 기질과 함께 진균성 생물막으로 성숙되는 과정은 최소 24-34시간이 소요된다[12]. 이에 본 연구에서는 진균 배양 후 5일까지 진균성 생물막 형성을 확인하였다. A. fumigatus 포자를 기관지 상피세포 혹은 비강상피세포와 함께 배양하는 경우 진균성 생물막의 형성이 증가된다[11,20]. 진균 포자들이 호흡상피세포 표면에 부착된 후 균사의 증식과 함께 세포외 기질이 형성되면서 진균 군집이 형성되고 생물막의 구멍(pore)과 내부 채널을 형성하면서 진균 생물막이 상피세포를 덮게 된다. 본 연구에서도 기존 연구와 마찬가지로 A. fumigatus와 비강상피세포를 함께 배양하는 경우 A. fumigatus 단독 배양의 경우보다 생물막의 형성이 배양 5일까지 지속적으로 증가됨을 확인할 수 있었다. 만성부비동염 환자의 비강 점막에서 진균성 생물막과 세균성 생물막이 공존함과 동시에 비강 내 세균에 의한 상피세포의 손상이 진균성 생물막의 형성을 촉진시킬 수 있다[7,8]. 본 연구에서도 LPS와 진균을 함께 배양한 경우 A. fumigatus 단독 혹은 비강상피세포와 A. fumigatus 배양한 경우 보다 많은 양의 진균성생물막이 형성됨을 확인할 수 있었다. 기존 연구의 경우 Staphylococcus [8]나 Pseudomonas [21]와 같은 균종과 진균을 함께 배양하는 경우 비부비동 점막의 선천성 면역방어기전의 손상 유도와 점막염증반응의 악화를 통해 생물막 형성이 이루어짐과 달리 본 연구의 경우 그람음성균 내독소인 LPS를 이용하여 진균 생물막 형성에 미치는 영향을 확인하였다. LPS의 경우 또한 균종과 마찬가지로 진균 생물막 형성에 영향을 미쳤는데 이는 LPS 또한 기존의 균종과 마찬가지로 상피세포의 손상을 직접 유도하거나, LPS가 비강상피세포의 세포외 기질 형성을 증가시키는 TGF-β1의 생성을 증가시킴으로써 생물막 형성을 유도할 수 있을 것으로 본다. 하지만 공초점 레이저 현미경의 경우 세포외 기질을 염색하는 concanavalin A의 형광강도가 배양 5일째 감소하였는데 이는 LPS와 진균의 합동배양 기간동안 배양액을 보충하거나 교체하지 않아 세포외 기질 형성을 위한 충분 조건을 만족시키지 못함에 기인할 것이다. 본 연구의 경우 체외 연구로 진행되어 인체 비강점막에서 일어나는 상피세포뿐 아니라 다양한 구조세포와 면역세포의 상호작용을 반영할 수 없어 진균성 생물막 형성을 충분히 이해하고 설명하기 어려운 한계점을 가지고 있다. 진균성 생물막의 형성과 관련된 동물 모델을 이용한 추가적인 연구가 필요할 것이다.
이상의 결과를 종합해 보면 비강상피세포를 A. fumigatus, LPS 단독 혹은 A. fumigatus와 LPS를 함께 배양하는 경우 IL-6, IL-8, TGF-β1 등의 화학매개물질 생성이 유의하게 증가함을 확인하였으나 A. fumigatus와 LPS를 함께 처리하거나 단독 처리한 경우 생성되는 화학매개물질의 양에는 차이를 보이지 않았다. 비강상피세포와 A. fumigatus를 함께 배양하는 경우 진균성 생물막의 형성이 증가하였으며, A. fumigatus와 LPS를 함께 배양하는 경우 진균성 생물막의 형성이 더욱 강하게 진행되었다. 본 연구를 통해 호흡기 염증성 질환의 경우 호흡기 내 존재하는 세균과 진균의 상호작용으로 생물막 형성이 활발하게 이루어지며, 이로 인해 염증반응이 유지 악화될 수 있음을 확인할 수 있었다.
Acknowledgements
This work was supported by the National Research Foundation of Korea (NRF) grant funded by the Korea government (Ministry of Science and ICT) (2019R1F1A1047757).
Notes
Author Contribution
Data curation: Tae-Gyun Kim, Chang-Ho WhangBo. Writing—original draft: Seung-Heon Shin. Writing—review & editing: JaeHo Yoo, Mi-Kyung Ye.