žĄú Ž°†
ž†źžē°žĚÄ ŪėłŪĚ°Íłį žÉĀŪĒľžĄłŪŹ¨žĚė ŪĎúŽ©īžĚĄ ŽćģÍ≥† žěąŽäĒ Ž¨ľžßąŽ°ú žäĶŽŹĄž°įž†ą ŽįŹ žú§Ūôú žěĎžö©Í≥ľ Ūē®ÍĽė žôłŽ∂ÄŽ°úŽ∂ÄŪĄį žú†žěÖŽźėŽäĒ Ž≥Ďžõźž≤ī(patho-gen)žôÄ ÍįĀžĘÖ ŪôĒŪēôŽ¨ľžßąžóź ŽĆÄŪēī Ž¨ľŽ¶¨ž†Ā Žį©žĖīŽ≤Ĺ žó≠Ūē†žĚĄ ŪēúŽč§[1]. ž†źžē° Žāīžóź Žč§žĖĎŪēú ŪėēŪÉúŽ°ú ž°īžě¨ŪēėŽäĒ ž†źžÜĆ(mucin)ŽäĒ ž†źžē°žĚė žĄĪžÉĀžĚĄ Í≤įž†ēŪēėŽäĒ ž§ĎžöĒŪēú ŽčĻŽč®ŽįĪŽ¨ľžßąŽ°ú ž†źžē°žú†ž†Ąžěźžóź žĚėŪēī ŽįúŪėĄžĚī ž°įž†ąŽźúŽč§[2]. ŪėłŪĚ°Íłį ž†źŽßČžóźŽäĒ 5ÍįÄžßÄ žĘÖŽ•ėžĚė ž£ľžöĒ ž†źžÜĆÍįÄ ŽĎź žłĶžúľŽ°ú ŽāėŽČėžĖīž†ł ž°īžě¨ŪēėŽ©į, ŽįĒÍĻ•žłĶžóźŽäĒ Ž∂ĄŽĻĄŪėē ž†źžÜƞ̳ MUC5ACžôÄ MUC5BÍįÄ, žēąžłĶžóźŽäĒ ŽßČÍ≤įŪē©Ūėē ž†źžÜƞ̳ MUC1, MUC4, MUC16žĚī ŪėłŪĚ°Íłį žÉĀŪĒľžĚė ž≤®Žč®Í≥ľ žĄ¨Ž™®žóź Ž∂Äžį©ŽźėžĖī ž°īžě¨ŪēúŽč§[3]. 5ÍįÄžßÄ ž£ľžöĒ ž†źžÜƎ吏ĚÄ Žč§žĖĎŪēú žěźÍ∑Ļžóź žĚėŪēī žĄúŽ°ú Žč§Ž•ł Ž∂ĄŽĻĄžĖĎžÉĀžĚĄ ŽāėŪÉÄŽāīŽ©į, žĚīŽäĒ Žč§žĖĎŪēú ŪėłŪĚ°Íłį žßąŪôėžĚė Ž≥ĎŪÉúžÉĚŽ¶¨žóź ŽįÄž†ĎŪēú ÍīÄŽ†®žĚī žěąŽäĒ Í≤ÉžúľŽ°ú žēĆŽ†§ž†ł žěąŽč§[3,4]. Í∑łŽü¨Žāė, ÍįúŽ≥Ąž†ĀžĚł ž†źžÜĆžĚė žÉĚŽ¶¨ž†ĀžĚł ÍłįŽä•žĚÄ ž∂©Ž∂ĄŪěą žēĆŽ†§ž†ł žěąžßÄ žēäžēĄ ž∂ĒÍįÄž†ĀžĚł žóįÍĶ¨ÍįÄ ŪēĄžöĒŪēú žč§ž†ēžĚīŽč§.
Ūô©žā¨(Asian sand dust)ŽäĒ ž£ľŽ°ú ŽīĄž≤† žēĄžčúžēĄ ŽĆÄŽ•ôžĚė ÍĪīž°įžßÄŽĆĞ󟞥ú Ž∂ąŽ†§ žė¨ŽĚľÍįĄ Žč§ŽüČžĚė ŪĚôŽ®ľžßÄÍįÄ žě•ÍĪįŽ¶¨ žĚīŽŹôŪēėžó¨ ŪēėŽäėžĚĄ ŽĖ†Žč§ŽčąŽ©į žĄúžĄúŪěą ŪēėÍįēŪēėŽäĒ ŪėĄžÉĀžĚīŽč§. Ūô©žā¨ŽäĒ žĚīžāįŪôĒÍ∑úžÜĆ(SiO2), žāįŪôĒžēĆŽ£®ŽĮłŽäĄ(Al2O3), žāįŪôĒž≤†(Fe2O3), žāįŪôĒžĻľžäė(CaO), žāįŪôĒžĻľŽ•®(K2O) ŽďĪ žēĹ 13žĘÖžĚė ÍłąžÜć ŽįŹ ŽĻĄÍłąžÜ枥Ī žāįŪôĒŽ¨ľÍ≥ľ ŽĮłŽüČžĚė ÍĶ¨Ž¶¨(Cu), ŪĀ¨Ž°¨(Cr), Žā©(Pb), žĻīŽďúŽģī(Cd), Žčąžľą(Ni) ŽďĪ 40žĘÖ žĚīžÉĀžĚė ŽĮłžĄłÍłąžÜćžě֞쟎°ú žĚīŽ£®žĖīž†ł žěąžúľŽ©į, ŽĆÄÍłį ž§Ďžóź ž°īžě¨ŪēėŽäĒ ŽāīŽŹÖžÜĆ(endotoxin)žôÄ ÍįôžĚÄ Žč§žĖĎŪēú žÉĚŽ¨ľžóźžĄú ÍłįžõźŪēú ŽĮłžĄł žě֞쟎ď§Í≥ľ Ūē®ÍĽė ŪĚ°žěÖŽźúŽč§[5]. Ž≥ĶŪē©Ž¨ľ ŪėēŪÉúžĚė Ūô©žā¨ŽäĒ žĶúÍ∑ľ žóįÍĶ¨žóźžĄú Žč§žĖĎŪēú Íłįž†ĄžĚĄ ŪÜĶŪēī ŪėłŪĚ°Íłį žóľž¶ĚŽįėžĚϞ̥ žú†ŽįúŪēėŽäĒ Í≤ÉžúľŽ°ú Ž≥īÍ≥†ŽźėÍ≥† žěąžúľŽ©į[6-9], ŪäĻŪěą ž†źžē°Ž∂ĄŽĻĄžôÄ ÍīÄŽ†®Žźú žóįÍĶ¨žóźžĄúŽäĒ interleukin-8Í≥ľ granulocyte-macrophage colony stimulating factoržôÄ ÍįôžĚÄ žóľž¶ĚžĄĪ žā¨žĚīŪ܆žĻīžĚł(cytokine)žĚī ž†źžē°žú†ž†Ąžěź Í≥ľŽįúŪėĄžóź žėĀŪĖ•žĚĄ ž£ľŽäĒ Í≤ÉžúľŽ°ú žēĆŽ†§ž†ł žěąžúľŽ©į[6], ŪäĻŪěą žĶúÍ∑ľ žóįÍĶ¨žóźžĄú Ūô©žā¨ÍįÄ ž£ľžöĒ Ž∂ĄŽĻĄŪėē ŪėłŪĚ°Íłį ž†źžÜƞ̳ MUC5BžôÄ MUC8žĚė Í≥ľŽ∂ĄŽĻĄŽ•ľ žú†ŽįúŪē®žĚī ŽįĚŪėÄž°ĆŽč§. ŪēėžßÄŽßĆ žā¨ŽěĆ ŪėłŪĚ°Íłį žÉĀŪĒľžĄłŪŹ¨žóźžĄú Ūô©žā¨ÍįÄ ž†źžē°žú†ž†Ąžěź ŽįúŪėĄžóź ŽĮłžĻėŽäĒ žėĀŪĖ•Í≥ľ Íłįž†Ąžóź ÍīÄŪēú žóįÍĶ¨ŽäĒ Žß§žöį ž†ĀžúľŽ©į, ŪäĻŪěą ŽßČÍ≤įŪē©Ūėē ž†źžÜĆžĚė ŽįúŪėĄžóź ŽĮłžĻėŽäĒ žėĀŪĖ•žóź ÍīÄŽ†®Žźú žóįÍĶ¨ŽäĒ ž†ĄŽ¨īŪēú žč§ž†ēžĚīŽč§.
ŽĒįŽĚľžĄú Ž≥ł žóįÍĶ¨žóźžĄúŽäĒ žā¨ŽěĆŪėłŪĚ°Íłį žÉĀŪĒľžĄłŪŹ¨žóźžĄú Ūô©žā¨ÍįÄ ŽĆÄŪĎúž†ĀžĚł ŽßČÍ≤įŪē©Ūėē ž†źžÜƞ̳ MUC4žôÄ MUC16 ŽįúŪėĄžóź ŽĮłžĻėŽäĒ žėĀŪĖ•Í≥ľ žč†Ūėłž†ĄŽč¨ Í≤ĹŽ°úŽ•ľ žēĆžēĄŽ≥īÍ≥†žěź žóįÍĶ¨Ž•ľ žčúŪĖČŪēėžėÄŽč§.
žě¨Ž£Ć ŽįŹ Žį©Ž≤ē
žě¨ Ž£Ć
žč§ŪóėžóźžĄú žā¨žö©Žźú Ūô©žā¨ŽäĒ ŽĆÄÍĶ¨ÍįÄŪÜ®Ž¶≠ŽĆÄŪēôÍĶź žĚėÍ≥ľŽĆÄŪēô žĚīŽĻĄžĚłŪõĄÍ≥ľÍĶźžč§žóźžĄú ž†úÍ≥ĶŽįõžēėŽč§. Ūô©žā¨ žÉėŪĒĆžĚė žĪĄž∑® Žį©Ž≤ēžĚĄ ÍįĄŽěĶŪēėÍ≤Ć žĄ§Ž™ÖŪēėŽ©ī, Ūô©žā¨ŽäĒ Ūô©žā¨ŪėĄžÉĀžĚī ŽįúžÉĚŪēėŽäĒ žčúÍłįžóź žĚłž≤úžóźžĄú ŽĆÄŽüČ ŪĚ°žěÖ Í≥ĶÍłį ŪŹ¨žßĎÍłįžĚł HV-500F(Sibata, Simata, Japan)Ž•ľ žĚīžö©Ūēėžó¨ žąėžßĎŪēėžėÄÍ≥†, ŪŹ¨žßĎÍłįžĚė žó¨Í≥ľžßÄŽ•ľ phosphate buffered saline(PBS) 10 mLžóź Ūúėž†ÄžĖī Ūô©žā¨ Ž∂ĄžßĄžĚĄ žĖĽžóąžúľŽ©į, žĖĽžĖīžßĄ žÉėŪĒƞ̥ 10 őľM ÍĶ¨Ž©ć ŪĀ¨ÍłįžĚė ŪēĄŪĄįŽ•ľ žā¨žö©Ūēėžó¨ žßÄŽ¶Ą 10 őľM žĚīŪēėžĚė ŪĀ¨ÍłįŽ•ľ ÍįÄžßÄŽäĒ Ūô©žā¨ žě֞쟎•ľ žĖĽžóąŽč§. žĚīŽ†áÍ≤Ć žĖĽžĖīžßĄ Ūô©žā¨ŽäĒ ž∂ĒÍįÄž†ĀžúľŽ°ú 121‚ĄÉžóźžĄú 15Ž∂Ą ŽŹôžēą ÍįÄžēēž¶ĚÍłįŽ©łÍ∑†(autoclave)žĚĄ Ūēėžó¨ Ž©łÍ∑†Žźú žÉĀŪÉúŽ°ú ŽßĆŽď§Í≥† -20‚ĄÉžóźžĄú ŽÉČŽŹô Ž≥īÍīÄŪēėžėÄŽč§ÍįÄ žč§Ūóėžóź žā¨žö©ŪēėžėÄŽč§[6]. Fetal bovine serum(FBS)žĚÄ HyClone Laboratories Inc.(Logan, UT, USA)žóźžĄú ÍĶ¨žěÖŪēėžėÄŽč§. Enhanced chemiluminescence kitsŽäĒ Perkin Elmer Lift Sciences(Boston, MA, USA)žóźžĄú ÍĶ¨žěÖŪēėžėÄžúľŽ©į, MUC4žĚė žĚľžį®Ūē≠ž≤īŽäĒ Neomarker(Zymed, San Francisco, CA, USA)žóźžĄú ÍĶ¨žěÖŪēėžėÄÍ≥†, anti-rabbit or anti-mouse horseradish peroxidase(HRP)-conjugated žĚīžį®Ūē≠ž≤īŽäĒ Santa Cruz Biotechnology(Santa Cruz, CA, USA)žóźžĄú ÍĶ¨žěÖŪēėžėÄŽč§.
Extracellular signal-regulated kinase(ERK)1/2 mitogenactivated protein kinase(MAPK), phospho-ERK1/2 MAPK, p38 MAPK ŽįŹ phospho-p38 MAPKŽäĒ cell signaling technology(Danvers, MA, USA)žóźžĄú ÍĶ¨žěÖŪēėžėÄžúľŽ©į, U0126(ERK1/2 MAPK žĄ†ŪÉĚž†Ā žĖĶž†úž†ú)žĚÄ Calbiochem(San Diego, CA, USA)žóźžĄú, SB203580(p38 MAPK žĄ†ŪÉĚž†Ā žĖĶž†úž†ú)žĚÄ BIOMOL(Plymouth Meeting, PA, USA)žóźžĄú ÍĶ¨žěÖŪēėžėÄŽč§. Toll-like receptor(TLR)4 small interfering ribonucleic acid(siRNA)žôÄ ERK1 MAPK siRNA, ERK2 MAPK siRNA, p38 MAPK siRNA, ŽĆÄž°įÍĶį siRNA, RPMI 1640 ŽįįžßÄ, OPTI-MEN I reduced Serum Medium, Lipofectamine 2000žĚÄ Invitrogen Corporation(Carlsbad, CA, USA)žóźžĄú ÍĶ¨žěÖŪēėžėÄÍ≥†, Human Keratinocyte Growth Supplement(HKGS, 5/500 mL of medium)ÍįÄ ž≤®ÍįÄŽźú EpiLife MediumžĚÄ Cascade Biologics(Portland, OR, USA)žóźžĄú ÍĶ¨žěÖŪēėžėÄŽč§.
žĄłŪŹ¨ ŽįįžĖĎ ŽįŹ ž≤ėžĻė
žā¨ŽěĆ ŪŹźžĚė ž†źžē°žÉĀŪĒľžĖĎ žēĒ žĄłŪŹ¨ž£ľ(human pulmonary mucoepidermoid carcinoma cell line)žĚł NCI-H292 žĄłŪŹ¨(American Type Culture Collection, Manassas, VA, USA)Ž•ľ 6-well platežóź 1√ó106 cells/wellžĚė ŽÜ掏ĄŽ°ú ž†ĎžĘÖŪēú ŪõĄ, 2 mM L-glutamine, 100 U/mL penicillin, 100 őľg/mL streptomycinÍ≥ľ 10% FBSÍįÄ ŪŹ¨Ūē®Žźú RPMI 1640 ŽįįžßÄŽ•ľ žĚīžö©Ūēėžó¨ 95%žĚė žāįžÜĆžôÄ 5%žĚė žĚīžāįŪôĒŪÉĄžÜĆÍįÄ ŪėľŪē©Žźú ŽįįžĖĎÍłįžóźžĄú 37‚ĄÉžĚė žė®ŽŹĄŽ°ú ŽįįžĖĎŪēėžėÄŽč§. 70~80% ž†ēŽŹĄžĚė žúĶŪē©ÍłįÍįÄ ŽźėŽ©ī žĄłŪŹ¨Ž•ľ 0.5% fetal calf serumžĚī ŪŹ¨Ūē®Žźú RPMI 1640 ŽįįžßÄŽ°ú ÍĶźž≤īŪēú ŪõĄ 24žčúÍįĄ ŽŹôžēą ŽįįžĖĎŪēėÍ≥†, Žč§žčú FBSÍįÄ ŪŹ¨Ūē®ŽźėžßÄ žēäžĚÄ RPMI 1640 ŽįįžßÄŽ°ú žĄłž≤ôŪēú ŪõĄ žč§Ūóėžóź žā¨žö©ŪēėžėÄŽč§.
Ūô©žā¨žĚė Ūö®Í≥ľŽ•ľ žēĆžēĄŽ≥īÍłį žúĄŪēīžĄú NCI-H292 žĄłŪŹ¨žóź 10 őľg/mLžôÄ 20 őľg/mL, 40 őľg/mL, 80 őľg/mL ŽÜ掏ĄžĚė Ūô©žā¨Ž•ľ ÍįĀÍįĀ Ūą¨žó¨ ŪõĄ ŽįįžĖĎŪēėžėÄŽč§. Í∑łŽ¶¨Í≥† U0126Í≥ľ SB203580žóź žĚėŪēú Ūô©žā¨žĚė Ūö®Í≥ľŽ•ľ žēĆžēĄŽ≥īÍłį žúĄŪēī NCI-H292 žĄłŪŹ¨žóź 10 őľg/mLžôÄ 20 őľg/mL, 40 őľg/mL, 80 őľg/mL ŽÜ掏ĄžĚė Ūô©žā¨Ž•ľ ž†Ą ž≤ėžĻėŪēėÍłį 1žčúÍįĄ ž†Ąžóź 2 őľMžĚė U0126, 20 őľMžĚė SB203580žĚĄ ÍįĀÍįĀ Ūą¨žó¨Ūēėžó¨ ŽįįžĖĎŪēėžėÄŽč§. ŽĆÄž°įÍĶįžĚÄ ŽŹôžĚľŪēú žčúÍįĄ ŽŹôžēą ŽįįžßĞ󟞥ú NCI-H292 žĄłŪŹ¨Ž•ľ Žč®ŽŹÖ ŽįįžĖĎŪēėžėÄŽč§.
žā¨ŽěĆžĚė ŪėłŪĚ°Íłį žÉĀŪĒľžĄłŪŹ¨Ž•ľ žĖĽÍłį žúĄŪēī žēĆŽ†ąŽ•īÍłįžóź ŽĆÄŪēú Íłįž†ÄžßąŪôėÍ≥ľ ÍįÄž°ĪŽ†•žĚī žóÜÍ≥†, ŪĒľŽ∂Ď讞쟞č§Ūóė(skin prick test)Í≥ľ multiple simultaneous allergen testžóźžĄú žĚĆžĄĪŽįėžĚĎžĚī Žāėžė® 10Ž™ÖžĚĄ ŽĆÄžÉĀžúľŽ°ú Ūēėžó¨ žúĶŽĻĄžą† žčúŪĖČ ž§Ďžóź ŪēėŽĻĄÍįĎÍįú ž°įžßĀžĚĄ žĖĽžóąŽč§. žĚľžį®ŽįįžĖϞ̥ ŪēėÍłį žúĄŪēī ŪēėŽĻĄÍįĎÍįú ž†źŽßČž°įžßĀžĚĄ PBSŽ°ú žĄłž≤ôŪēú ŪõĄ, 90Ž∂Ą ŽŹôžēą ŪēėŽĻĄÍįĎÍįú ž†źŽßČž°įžßĀžĚī žĻ®žąėŽź† ž†ēŽŹĄŽ°ú dispase(Boehringer Mannheim Biochemica, Mannheim, Germany)žóź žĻ®ž†ĄžčúžľįŽč§. žĚīŪõĄ 15Ž≤ą žąėžą†žö© žĻľŽ°ú ŪēėŽĻĄÍįĎÍįú ž†źŽßČž°įžßĀžĚĄ ÍĻ®ŽĀóŪēėÍ≤Ć ÍłĀžĖīŽāīžĖī 1% PBSžóź ŽčīžĚÄ ŪõĄ, 200 meshžóź ÍĪłŽü¨ ŽāīÍ≥† ŪäúŽłĆžóź ŽčīžēĄ 3000 rpmžóźžĄú 5Ž∂ĄÍįĄ žõźžč¨ Ž∂ĄŽ¶¨ŪēėžėÄŽč§. žÉĀžłĶŽ∂ÄžĚė PBSŽ•ľ ž†úÍĪįŪēėÍ≥† HKGSÍįÄ ž≤®ÍįÄŽźú EpiLife Medium 6 mLŽ•ľ ŽĄ£žĚÄ ŪõĄ 24-well platežóź 1 mLžĒ© ž†ĎžĘÖŪēėžėÄŽč§. ŪėĄŽĮłÍ≤ĹžúľŽ°ú žĄłŪŹ¨žĚė žĖϞ̥ ŪôēžĚłŪēú ŪõĄ 2žĚľŽßąŽč§ HKGSŽ•ľ ž≤®ÍįÄŪēú EpiLife MediumžúľŽ°ú ŽįįžßÄŽ•ľ ÍĶźž≤īŪēėŽ©īžĄú žĄłŪŹ¨ÍįÄ 70~80% ž†ēŽŹĄŽ°ú ž¶ĚžčĚŽźėŽ©ī žč§Ūóėžóź žā¨žö©ŪēėžėÄŽč§. žĚľžį® ŽįįžĖĎŪēú ŪēėŽĻĄÍįĎÍįú ž†źŽßČž°įžßĀžóźžĄú Ūô©žā¨žĚė Ūö®Í≥ľŽ•ľ žēĆžēĄŽ≥īÍłį žúĄŪēīžĄú Ūô©žā¨ 40 őľg/mLŽ•ľ ž†Ą ž≤ėžĻėŪēú ŪõĄ 1žčúÍįĄ Ží§žóź ÍįĀÍįĀ 2 őľMžĚė U0126, 20őľMžĚė SB203580žĚĄ Ūą¨žó¨Ūēėžó¨ ŽįįžĖĎŪēėžėÄŽč§. ŽĆÄž°įÍĶįžĚÄ ŽŹôžĚľŪēú žčúÍįĄ ŽŹôžēą ŽįįžßĞ󟞥ú ŪėłŪĚ°Íłį žÉĀŪĒľžĄłŪŹ¨žôÄ NCI-H292 žĄłŪŹ¨Ž•ľ ÍįĀÍįĀ Žč®ŽŹÖžúľŽ°ú ŽįįžĖĎŪēėžėÄŽč§. žč§ŪóėžóźžĄú žā¨žö©Ūēú NCI-H292 žĄłŪŹ¨žôÄ žĚľžį® ŽįįžĖĎŽźú ŪēėŽĻĄÍįĎÍįú ž†źŽßČ žÉĀŪĒľžĄłŪŹ¨žóź ŽĆÄŪēú Ūô©žā¨žĚė žĄłŪŹ¨ŽŹÖžĄĪ žú†Ž¨īŽäĒ MTT Ž∂ĄžĄĚ(Sigma-Aldrich, St. Louis, MO, USA)Í≥ľ ŪėĄŽĮłÍ≤Ş̥ žĚīžö©Ūēú žĄłŪŹ¨ ŪėēŪÉú Ž≥ÄŪôĒ žú†Ž¨īŽ•ľ ŪôēžĚłŪēėžó¨ Í≤Äž¶ĚŪēėžėÄŽč§.
Ž≥ł žóįÍĶ¨ŽäĒ Ž≥łžõź žěĄžÉĀžčúŪóėžč¨žā¨žúĄžõźŪöĆ(Institutional Review Board)žĚė žäĻžĚłžĚĄ ŽįõžēĄ žčúŪĖČŪēėžėÄŽč§.
Reverse transcription polymerase chain reaction(RT-PCR) Ž∂ĄžĄĚ
PTC-200(MF Reseach Inc., Watertown, MA, USA) polymerase chain reaction(PCR) machineÍ≥ľ Gene Amp RNA PCR core kit(Applied Biosystems, Foster City, CA, USA)Ž•ľ žā¨žö©Ūēėžó¨ ž†úž°įžā¨žĚė Žį©Ž≤ēŽĆÄŽ°ú žčúŪĖČŪēėžėÄŽč§. PCRžóź žā¨žö©Žźú oligonucleotide primerŽäĒ ŽįĚŪėĞߥ žóľÍłįžĄúžóīžóź žĚėŪēī ž†úžěĎŽźėžóąžúľŽ©į, ÍįĀ ŽįėžĚĎžĚė ŽāīŽ∂Ä žĖĎžĄĪ ŽĆÄž°įÍĶį(internal positive control)žĚÄ glyceraldehyde-3-phosphate dehydrogenase(GAPDH)Ž•ľ žā¨žö©ŪēėžėÄŽč§. žč§Ūóėžóź žā¨žö©Žźú primeržĚė žóľÍłįŽįįžóīžĚÄ MUC4žĚė Í≤Ĺžöį senseŽäĒ 5‚Äô-TTC TAA GAA CCA CCA GAC TCA GAG C-3‚Äô, antisenseŽäĒ 5‚Äô-GAG ACA CAC CTG CAG AGA ATG AGC-3‚ÄôžĚīŽč§. MUC16žĚė Í≤Ĺžöį senseŽäĒ 5‚Äô-GGC TCT ACC TTA ACG GTT ACA ATG AA-3‚Äô, antisenseŽäĒ 5‚Äô-GGT ACC CCA TGG CTG TTG TG-3‚ÄôžĚīŽč§. GAPDHžĚė Í≤Ĺžöį senseŽäĒ 5‚ÄôCCT CCA AGG AGT AAG ACC CC-3‚Äô, antisenseŽäĒ 5‚Äô-AGG GGT CTA CAT GGC AAC TG-3‚ÄôžĚīžóąŽč§. žč§Ūóėžóź žā¨žö©ŽźėžĖī ž¶ĚŪŹ≠Žźú mRNA žāįŽ¨ľžĚė ŪĀ¨ÍłįŽäĒ MUC4ŽäĒ 467 bp, MUC16žĚÄ 114 bp, GAPDHŽäĒ 145 bpžėÄŽč§. MUC4, MUC16žĚė reverse transcription polymerase chain reaction(RT-PCR)žĚė ž°įÍĪīÍ≥ľ Í≥ľž†ēžĚÄ žĚīŽĮł Ž≥īÍ≥†Žźú žóįÍĶ¨Žď§žĚė Žį©žčĚŽĆÄŽ°ú žčúŪĖČŪēėžėÄŽč§[10]. ž¶ĚŪŹ≠Žźú ž§ĎŪē©Ūö®žÜĆžóįžáĄŽįėžĚĎžĚė žāįŽ¨ľžĚÄ SYBR greenžĚī Ūē®žú†Žźú 1% agarose gelžĚĄ ŪÜĶŪēú ž†ĄÍłįžėĀŽŹôžĚĄ žĚīžö©Ūēėžó¨ Ž∂ĄŽ¶¨ ÍīÄžįįŪēėžėÄŽč§. ŪôēžĚłŽźú ŽĚ†(band)žĚė žĄłÍłįŽäĒ Scion Image software(Scion Corporation, Frederick, MD, USA)Ž•ľ žĚīžö©Ūēėžó¨ Žįėž†ēŽüČž†ĀžúľŽ°ú Ž∂ĄžĄĚŪēėžėÄÍ≥†, ŽĆÄž°įÍĶįžĚė densityŽ•ľ 100žúľŽ°ú ŪēėžėĞ̥ ŽēĆ žč§ŪóėÍĶįžĚė density ÍįížĚĄ ŽĻĄžú®Ž°ú ŽāėŪÉÄŽāīžĖī relative densityŽ°ú ŽāėŪÉÄŽāīžóąŽč§.
Real-time PCR Ž∂ĄžĄĚ
Ūē©žĄĪŽźú cDNA 1 őľLŽ•ľ ŽĆÄžÉĀžúľŽ°ú LC Fast Start DNA Master SYBR Green kit(Roche Applied Science, Mannheim, Germany)Ž•ľ žā¨žö©Ūēėžó¨ real-time PCRžĚĄ žĚīžö©Ūēú ž†źžē°žú†ž†Ąžěź ŽįúŪėĄŽįėžĚϞ̥ žčúŪĖČŪēėžėÄŽč§. Real-time PCRžĚÄ žĶúžĘÖŽüČžĚī 10 őľLÍįÄ ŽźėÍ≤Ć 2.5 mMžĚė MgCl2žôÄ žĶúžĘÖ ŽÜ掏ĄÍįÄ 0.5 őľMžĚī ŽźėÍ≤Ć primerŽ•ľ Ūą¨žó¨ŪēėžėÄžúľŽ©į, 25 ngžĚė RNA 1 őľLŽ•ľ žĚīžö©Ūēėžó¨ žč§ŪóėžĚĄ žąėŪĖČŪēėžėÄŽč§. ž†ēŽüČž†ĀžĚł PCRžĚÄ Light-Cycler(Roche Applied Science, Mannheim, Germany)Ž•ľ žā¨žö©Ūēėžó¨ 95‚ĄÉžóźžĄú 10žīąÍįĄ Ž≥ÄžĄĪ(denaturation) Í≥ľž†ēžĚĄ ÍĪįžĻėÍ≥† 60‚ĄÉžóźžĄú 5žīąÍįĄ Í≤įŪē©(annealing) ŽįėžĚϞ̥ žčúŪā® ŪõĄ, 72‚ĄÉžóźžĄú 10žīąÍįĄ žóįžě•(extension) ŽįėžĚĎŪēėžėÄÍ≥†, žĚīŽü¨Ūēú Í≥ľž†ēžĚĄ 45ŪöĆ ŽįėŽ≥ĶŪēėžėÄŽč§. ž¶ĚŪŹ≠žĚė ž†ēŪôēŽŹĄŽäĒ Ž≥ÄŪėēÍ≥°žĄ†(melting curve; Roche Applied Science, Mannheim, Germany)žĚĄ žā¨žö©Ūēėžó¨ ŪŹČÍįÄŪēėžėÄŽč§.
Ž©īžó≠Ž∂ĄžĄĚŽ≤ē(Immunoassay)
MUC4 ŽčĻŽč®ŽįĪžĚė Ūē®ŽüȞ̥ žł°ž†ēŪēėÍłį žúĄŪēīžĄú enzyme-linked immunosorbent assay(ELISA)Ž≤ēžĚĄ žĚīžö©ŪēėžėÄŽč§. žčúŽ£ĆŽ•ľ ž≤ėŽ¶¨Ūēú ŽįįžĖĎŽźú žĄłŪŹ¨žóźžĄú lysis buffer[50 mM tris¬∑Cl(pH 7.5), 1 mM ethylene glycol tetraacetic acid, 1% triton X-100, and 1 mM phenylmethylsulfonyl fluoride]Ž°ú Žč®ŽįĪžĚĄ ž∂Ēž∂úŪēėžó¨ ž†ēŽüČŪēėžėÄŽč§. ž∂Ēž∂úŪēú Žč®ŽįĪ 100 őľgžĚĄ 96-well platežóź ŽčīÍ≥† 40‚ĄÉžóźžĄú ÍĪīž°įŽź† ŽēĆÍĻĆžßÄ Žį©žĻėŪēú ŪõĄ plateŽ•ľ PBSŽ°ú 3ŪöĆ žĄłž≤ôŪēėžėÄŽč§. ŽĻĄŪäĻžĚīž†Ā Í≤įŪē©žĚĄ Žį©žßÄŪēėÍłį žúĄŪēī 2% bovine serum albuminžúľŽ°ú žč§žė®žóźžĄú 1žčúÍįĄ ŽŹôžēą žį®Žč®Ūēú ŪõĄ PBSŽ°ú 3ŪöĆ žĄłž≤ôŪēú Žč§žĚĆ 0.05% tween 20žĚĄ Ūē®žú†Ūēú PBSžóź 1:200žúľŽ°ú Ų̄žĄĚŽźú MUC4 žĚľžį®Ūē≠ž≤īŽ°ú ŽįėžĚĎžčúžľįŽč§. Žč§žčú PBSŽ°ú 3ŪöĆ žĄłž≤ôŪēú ŪõĄ HRPconjugated žĚīžį®Ūē≠ž≤īŽ•ľ 0.05% tween 20žĚĄ Ūē®žú†Ūēú PBSžóź 1:5000žúľŽ°ú Ų̄žĄĚŪēėžó¨ ÍįĀ wellžóź ž≤®ÍįÄŪēėžėÄÍ≥†, 1žčúÍįĄ ŪõĄžóź ÍįĀ wellžĚĄ PBSŽ°ú 3ŪöĆ žĄłž≤ôŪēėžėÄŽč§. 3, 3‚Äô, 5, 5‚Äô-tetramethylbenzidine žö©žē°žúľŽ°ú ŽįúžÉČŪēú ŪõĄ, 2N-H2SO4Ž•ľ žĚīžö©Ūēėžó¨ ž§ĎŽč®žčúžľįŽč§. ELISA reader(EL800‚ďá; BIO-TEK Instruments, Winooski, VT, USA)Ž°ú 450 nmžóźžĄú ŪĚ°ÍīĎŽŹĄŽ•ľ žł°ž†ēŪēú ŪõĄ ŪĎúž§ÄÍ≥°žĄ†žĚĄ žĚīžö©Ūēėžó¨ Žč®ŽįĪžĚė žĖϞ̥ ž†ēŽüČŪēėžėÄŽč§.
Westen blot analysis of p38 and ERK1/2 MAPK phophorylation
NCI-H292 žĄłŪŹ¨Ž•ľ 6-well platežóź ŽčīÍ≥† ŽÜ掏Ąžóź ŽĒįŽĚľ Ūô©žā¨Ž•ľ ž≤ėŽ¶¨Ūēėžó¨ ž†ĎžĘÖ ŪõĄ ŽįįžĖĎŪēėžėÄŽč§. žĚīŪõĄ trypsinžóź ÍįĀ žĄłŪŹ¨Žď§žĚĄ ŽÖłž∂úžčúŪā® ŪõĄ lysis bufferŽ•ľ ÍĪįž≥ź 4‚ĄÉžóźžĄú 700√ógžúľŽ°ú žõźžč¨ Ž∂ĄŽ¶¨žčúžľú ÍįĀÍįĀžĚė pelletžĚĄ ŽßƎ吏óąŽč§. ÍįĀÍįĀžĚė pelletžĚĄ lysis bufferžóźžĄú Ž∂Äžú†Ūēú ŪõĄ žõźžč¨Ž∂ĄŽ¶¨Ž•ľ žčúŪĖČŪēėžó¨ ž†ēž†úŽźú žÉĀž∂©žē°žĚĄ whole-cell lysateŽ°ú Ž≥īÍīÄŪēėžėÄŽč§. žĚīŽü¨Ūēú Žį©Ž≤ēžúľŽ°ú Ž∂ĄŽ¶¨Žźú 50 őľgžĚė Žč®ŽįĪžĚĄ 10% reducing sodium dodecyl sulfatepolyacrylamide gelžĚĄ žĚīžö©Ūēú ž†ĄÍłįžėĀŽŹôžĚĄ žč§žčúŪēėžėÄŽč§. ž†ĄÍłį žėĀŽŹôŽźú√óŽč®ŽįĪžßąžĚĄ nitrocellulose membranežúľŽ°ú žėģÍ≤® tris-buffer salineÍ≥ľ tween-20 buffer[20 mM/L Tris-HCl(pH 7.6), 135 mM/L NaCl, 1% tween-20]žôÄ 5% nonfat dry milkŽ°ú ž≤ėŽ¶¨Ūēėžó¨ Ūē≠ž≤īžôÄžĚė ŽĻĄŪäĻžĚīž†Ā Í≤įŪē©žĚĄ žĖĶž†úžčúŪā® ŪõĄ ERK1/2 MAPKžôÄ p38 MAPK žĚľžį®Ūē≠ž≤īŽ°ú ÍįĀÍįĀ 4žčúÍįĄ ŽįėžĚĎžčúžľįŽč§. žĚīŪõĄ trisbuffered salinežôÄ tween-20 bufferŽ°ú žĄłž≤ôŪēėžėÄžúľŽ©į antimouse HRP-conjugated ERK1/2 MAPKžôÄ p38 MAPK žĚīžį®Ūē≠ž≤īŽ°ú ÍįĀÍįĀ 1žčúÍįĄ ŽįėžĚĎŪēú Žč§žĚĆ žĄłž≤ôŪēėžó¨ enhanced chemiluminescence reagent kitŽ•ľ žĚīžö©Ūēėžó¨ ŪėĄžÉĀŪēėÍ≥† 10žīąÍįĄ X-ray filmžóź ÍįźÍīĎŪēėžó¨ ÍįĀÍįĀžĚė ŽĚ†(band)Ž•ľ ŪôēžĚłŪēėžėÄŽč§. ÍįźÍīĎŽźú ŽĚ†žĚė žĄłÍłįŽäĒ Scion Image software(Scoin Corporation, Frederick, MD, USA)Ž•ľ žĚīžö©Ūēėžó¨ Žįėž†ēŽüČž†ĀžúľŽ°ú Ž∂ĄžĄĚŪēėžó¨ žÉĀŽĆÄž†ĀžĚł densityŽ°ú ŽāėŪÉÄŽāīžóąŽč§.
siRNA Ūėēžßą ž†ĄŪôė
siRNA Ūėēžßą ž†ĄŪôė žč§Ūóė žā¨žö© žąúžĄú ŽįŹ ž°įÍĪīžĚÄ ERK1, ERK2 ŽįŹ p38 MAPKžĚė siRNAžóź ŽĆÄŪēī Í≤ĆžčúŽźú ž†úž°įžā¨žĚė Žį©Ž≤ēŽĆÄŽ°ú žčúŪĖČŪēėžėÄŽč§(Invitrogen Corporation). ÍįĄŽěĶŪěą žĄ§Ž™ÖŪēėžěźŽ©ī NCIH292 žĄłŪŹ¨Ž•ľ 6-well platežóź 1√ó105 cells/wellžĚė ŽÜ掏ĄŽ°ú ž†ĎžĘÖ ŪõĄ, RPMI 1640 ŽįįžßÄŽ•ľ žĚīžö©Ūēėžó¨ Ūē≠žÉĚž†ú žóÜžĚī ŪēėŽ£ĽŽį§ ŽŹôžēą ŽįįžĖĎŪēėžėÄŽč§. Žč§žĚĆ Žā† 80~90% ž†ēŽŹĄžĚė žúĶŪē©ÍłįÍįÄ ŽźėžĖī PBSŽ°ú žĄłž≤ôŪēú ŪõĄ, OPTI-MEN I Reduced Serum MediumžĚĄ žĄłŪŹ¨žóź ž≤®ÍįÄŪēėžėÄŽč§. ERK1 MAPK siRNAžôÄ ŪēĶžāį ž†ĄžÜ° Ž¨ľžßąžĚł Lipofectamine 2000žĚĄ ERK1 MAPK siRNA-Lipofectamine Ž≥ĶŪē©ž≤īŽ•ľ ŽßĆŽď§Íłį žúĄŪēī 20Ž∂ĄÍįĄ OPTI-MEN I Reduced Serum MediumžóźžĄú ŽįįžĖĎ ŪõĄ, ERK1 MAPK siRNA-Lipofectamine Ž≥ĶŪē©ž≤īŽ•ľ Ūē®žú†Ūēú ŽįįžßÄŽ•ľ ERK1 MAPK siRNA 20 nM ŽÜ掏ĄŽ°ú ÍįĀÍįĀžĚė wellžóź ž≤®ÍįÄ ŪõĄ CO2 ŽįįžĖĎÍłįžóźžĄú 37‚ĄÉŽ°ú 24žčúÍįĄ ŽįįžĖĎŪēėžėÄŽč§. ERK1 MAPK siRNA-Lipofectamine Ž≥ĶŪē©ž≤ī ŽįįžßÄŽ•ľ Ūėēžßą ž†ĄŪôė ŪôúžĄĪžĚė žÜźžč§ žóÜžĚī 4žčúÍįĄ ŪõĄ RPMI I640 ŽįįžßÄŽ°ú ŽĆÄž≤īŪēú ŪõĄ ERK1 MAPK siRNAŽ•ľ žĚīžö©Ūēėžó¨ 24žčúÍįĄ ŽŹôžēą Ūėēžßą ž†ĄŪôėžĚĄ ÍĪįžĻú žĄłŪŹ¨Ž•ľ 10 ng/mL ŽÜ掏ĄžĚė Ūô©žā¨žóź ŽÖłž∂úžčúŪā§Í≥†, RT-PCRžĚĄ žĚīžö©Ūēėžó¨ MUC4 mRNAžĚė ŽįúŪėĄŽüȞ̥ žł°ž†ēŪēėžėÄŽč§. NCI-H292 žĄłŪŹ¨žóźžĄú ERK1 MAPK siRNAžĚė Ūėēžßą ž†ĄŪôėžĚÄ 90% žĚīžÉĀžúľŽ°ú žł°ž†ēŽźėžóąÍ≥†, TLR4 siRNAžôÄ ERK2 MAPK siRNA, p38 MAPK siRNA, ŽĆÄž°įÍĶį siRNAžĚė Ūėēžßą ž†ĄŪôėŽŹĄ žúĄžôÄ ŽŹôžĚľŪēú Í≥ľž†ēžúľŽ°ú žąėŪĖČŪēėžėÄŽč§.
ŪÜĶ Í≥Ą
Ž™®Žď† žč§ŪóėžĚÄ 3žį®Ž°Ä žĚīžÉĀžĚė ŽįėŽ≥Ķ žč§ŪóėžĚĄ ŪÜĶŪēī žĖĽžĚÄ Í≤įÍ≥ľÍįížĚĄ žĚīžö©Ūēėžó¨ Ž∂ĄžĄĚŪēėžėÄžúľŽ©į, ŪÜĶÍ≥Ą ž≤ėŽ¶¨ŽäĒ Windowsžö© SPSS version 10.0(SPSS Inc., Chicago, IL, USA)žĚĄ žā¨žö©ŪēėžėÄŽč§. Ž™®Žď† žč§ŪóėžĚÄ pÍįížĚī 0.05 ŽĮłŽßƞ̳ Í≤ĹžöįŽ•ľ žú†žĚėŪēú Í≤ÉžúľŽ°ú ž†ēŪēėžó¨ Student‚Äôs t-testžôÄ Mann-Whitney U testŽ•ľ žĚīžö©Ūēėžó¨ Ž∂ĄžĄĚŪēėžėÄŽč§.
Í≤į Í≥ľ
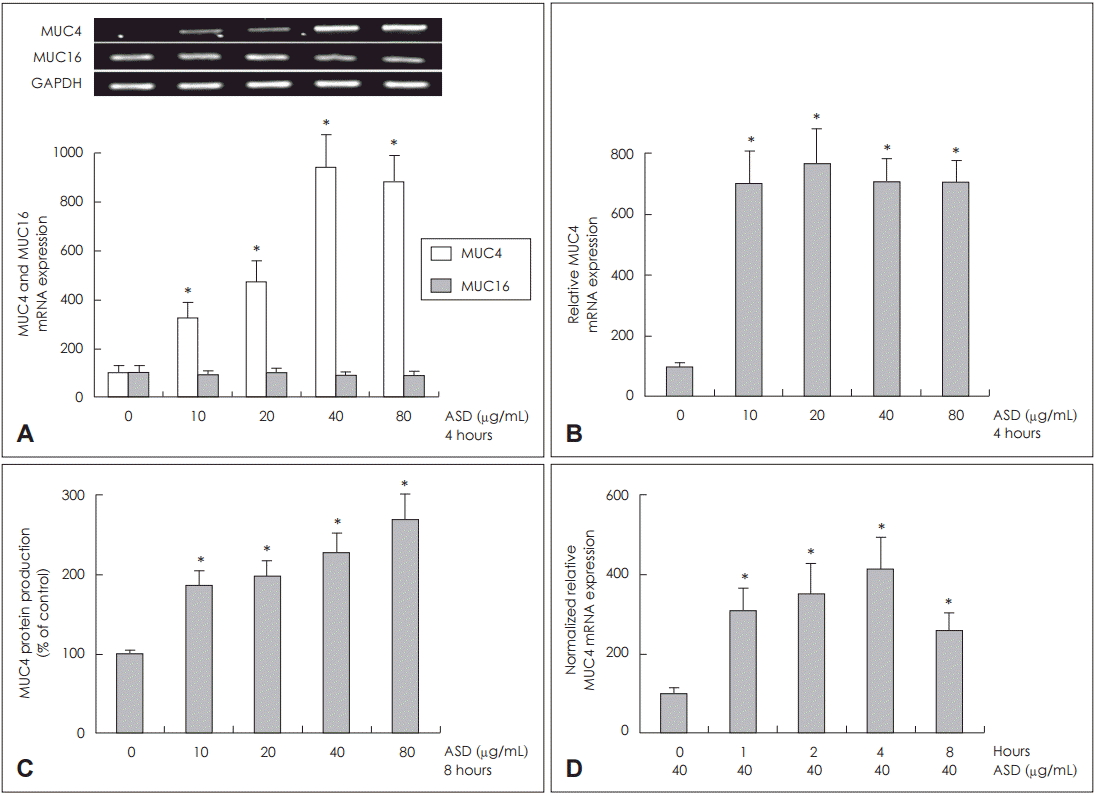
NCI-H292 žĄłŪŹ¨žóźžĄú Ūô©žā¨ÍįÄ MUC4žôÄ MUC16 ŽįúŪėĄžóź ŽĮłžĻėŽäĒ žėĀŪĖ•
Ūô©žā¨ÍįÄ ŽĮłžĻėŽäĒ ž£ľžöĒ ŪėłŪĚ°Íłį ŽßČÍ≤įŪē©ž†źžÜƞ̳ MUC4žôÄ MUC16 ŽįúŪėĄžĚĄ žēĆÍłį žúĄŪēīžĄú NCI-H292 žĄłŪŹ¨žóź ÍįĀÍįĀ Žč§Ž•ł ŽÜ掏ĄžĚė Ūô©žā¨Ž•ľ Ūą¨žó¨Ūēėžó¨ 4žčúÍįĄ ŽŹôžēą ŽįįžĖĎŪēėžėÄŽč§. RT-PCR Ž∂ĄžĄĚ Í≤įÍ≥ľ, Ūô©žā¨ÍįÄ Ž™®Žď† žö©ŽüȞ󟞥ú MUC4 mRNA ŽįúŪėĄžĚĄ žĚėŽĮł žěąÍ≤Ć ž¶ĚÍįÄŪēėžėÄžúľŽāė, MUC16 mRNA ŽįúŪėĄžĚÄ ŽĆÄž°įÍĶįÍ≥ľ žį®žĚīÍįÄ žóÜžóąŽč§(Fig. 1A). Ūô©žā¨žĚė žö©ŽüČžóź ŽĒįŽ•ł MUC4 mRNA ŽįúŪėĄÍ≥ľ ŽčĻŽč®ŽįĪ žÉĚžĄĪžĚĄ žēĆžēĄŽ≥īÍ≥†žěź NCI-H292 žĄłŪŹ¨žóź Ūô©žā¨Ž•ľ Ūą¨žó¨ŪēėÍ≥† ÍįĀÍįĀ 4žčúÍįĄÍ≥ľ 8žčúÍįĄ ŽŹôžēą ŽįįžĖĎŪēú ŪõĄ, žčúŪĖČŪēú real time PCRÍ≥ľ ELISAžĚė Ž∂ĄžĄĚ Í≤įÍ≥ľžóźžĄú MUC4 mRNA ŽįúŪėĄÍ≥ľ ŽčĻŽč®ŽįĪ žÉĚžĄĪžĚÄ Ž™®Žď† žö©ŽüȞ󟞥ú ž¶ĚÍįÄŪēėžėÄžúľŽāė, žö©ŽüČžĚėž°īžĄĪžĚÄ žóÜžóąŽč§(Fig. 1B and C).
Ūô©žā¨žôÄ ŽįįžĖĎ žčúÍįĄžóź ŽĒįŽ•ł MUC4 mRNA ŽįúŪėĄžĚĄ žēĆžēĄŽ≥īÍ≥†žěź Ūô©žā¨ 40 őľgžĚĄ NCI-H292 žĄłŪŹ¨žóź Ūą¨žó¨ŪēėÍ≥† ÍįĀÍįĀ 1žčúÍįĄ, 2žčúÍįĄ, 4žčúÍįĄ, 8žčúÍįĄ, 24žčúÍįĄžĚĄ ŽįįžĖĎŪēú ŪõĄ real time PCRŽ°ú MUC4 mRNA ŽįúŪėĄžĚĄ Ž∂ĄžĄĚŪēú Í≤įÍ≥ľ ŽįįžĖĎ 4žčúÍįĄ ŪõĄ MUC4 mRNA ŽįúŪėĄžĚī ÍįÄžě• ž¶ĚÍįÄŽźėžóąžúľŽāė, žčúÍįĄ žĚėž°īžĄĪžĚÄ ÍīÄžįįŽźėžßÄ žēäžēėŽč§(Fig. 1D).
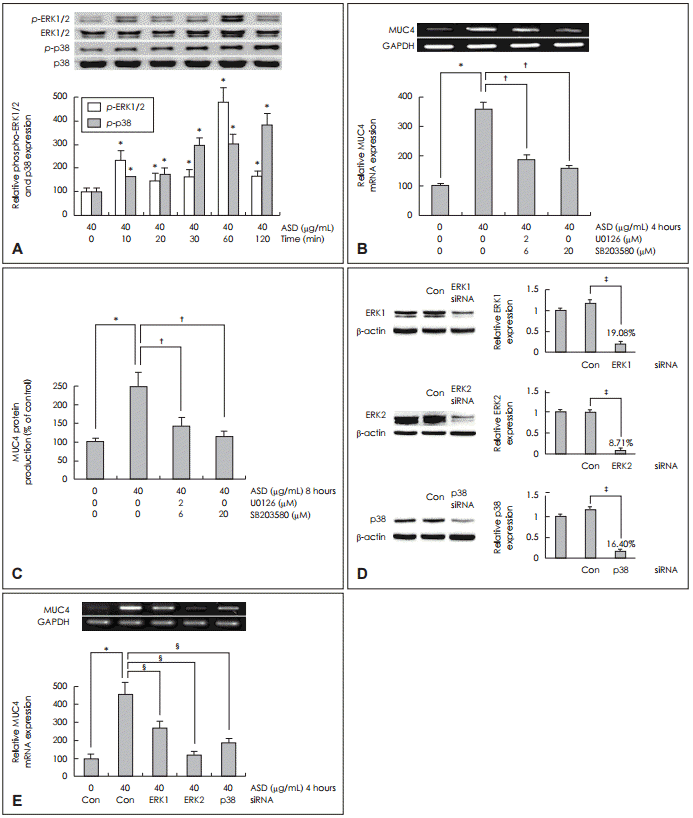
NCI-H292 žĄłŪŹ¨žóźžĄú Ūô©žā¨ÍįÄ ERK1/2 MAPKžôÄ p38 MAPKŽ•ľ ŪÜĶŪēú MUC4 ŽįúŪėĄžóź ŽĮłžĻėŽäĒ žėĀŪĖ•
Ūô©žā¨žóź žĚėŪēī žú†ŽŹĄŽźú MUC4 ŽįúŪėĄžóźžĄú žč†Ūėłž†ĄŽč¨ Í≤ĹŽ°úŽ•ľ žēĆžēĄŽ≥īÍłį žúĄŪēėžó¨ NCI-H292 žĄłŪŹ¨žóźžĄú ERK1/2 MAPKžôÄ p38 MAPK žč†Ūėłž†ĄŽč¨ Í≤ĹŽ°úŽ•ľ Ž∂ĄžĄĚŪēėžėÄŽč§. Ūô©žā¨ 40 őľgžĚĄ Ūą¨žó¨Ūēú ŪõĄ žčúÍįĄžóź ŽĒįŽ•ł ERK1/2 MAPKžôÄ p38 MAPKžĚė žĚłžāįŪôĒŽ•ľ Western blotžúľŽ°ú Ž∂ĄžĄĚŪēú Í≤įÍ≥ľžóźžĄú ERK1/2 MAPKžôÄ p38 MAPKÍįÄ žčúÍįĄžĚī Í≤ĹÍ≥ľŪē®žóź ŽĒįŽĚľ žĚłžāįŪôĒÍįÄ žĚėŽĮł žěąÍ≤Ć ž¶ĚÍįÄŪēėžėÄŽč§(Fig. 2A).
ERK1/2 MAPKžôÄ p38 MAPKžĚė žĚłžāįŪôĒÍįÄ Ūô©žā¨žóź žĚėŪēī žú†ŽŹĄŽźú MUC4 ŽįúŪėĄžĚĄ ž¶ĚÍįÄžčúŪā§ŽäĒ žč†Ūėłž†ĄŽč¨ Í≤ĹŽ°úŽ•ľ ŪôēžĚłŪēėÍłį žúĄŪēīžĄú NCI-H292 žĄłŪŹ¨žóź ERK1/2 MAPKžĚė žĖĶž†úž†úžĚł U0126(2 őľM)Í≥ľ p38žĚė žĖĶž†úž†úžĚł SB203580(20 őľM)žúľŽ°ú ž†Ą ž≤ėžĻė ŪõĄ Ūô©žā¨ 40 őľgžĚĄ Ūą¨žó¨ŪēėÍ≥† ÍįĀÍįĀ 4žčúÍįĄÍ≥ľ 8žčúÍįĄ ŽŹôžēą ŽįįžĖĎŪēėžėÄŽč§. RT-PCRÍ≥ľ ELISAŽ°ú Ž∂ĄžĄĚŪēú Í≤įÍ≥ľžóźžĄú Ūô©žā¨žóź ŽĒįŽ•ł MUC4 mRNA ŽįúŪėĄÍ≥ľ ŽčĻŽč®ŽįĪ žÉĚžĄĪžĚī ŽĆÄž°įÍĶįÍ≥ľ ŽĻĄÍĶźŪēėžėĞ̥ ŽēĆ žĚėŽĮłžěąÍ≤Ć ÍįźžÜĆŪēėžėÄŽč§(Fig. 2B and C).
ŽėźŪēú ERK1/2 MAPKžôÄ p38 MAPKžĚė žč†Ūėłž†ĄŽč¨ Í≤ĹŽ°úŽ•ľ Í≤Äž¶ĚŪēėÍłį žúĄŪēėžó¨ NCI-H292 žĄłŪŹ¨Ž•ľ ERK1 MAPK siRNAžôÄ ERK2 MAPK siRNA, p38 MAPK siRNAŽ°ú ÍįĀÍįĀ ŪĎúž†Ā žú†ž†ĄžěźŽ•ľ knockdownžčúŪā® ŪõĄ Western blotžúľŽ°ú Ž∂ĄžĄĚŪēėžėÄžúľŽ©į ž∂©Ž∂ĄŪēú Ūėēžßą ž†ĄŪôėžĚī žĚľžĖīŽā¨žĚƞ̥ ŪôēžĚłŪēėžėÄÍ≥†(Fig. 2D), ÍįĀÍįĀžĚė ŪĎúž†Āžú†ž†Ąžěź Ūėēžßą ž†ĄŪôėžĚĄ žčúŪĖČŪēú ŪõĄ Ūô©žā¨ 40 őľgžĚĄ Ūą¨žó¨ŪēėžėÄžúľŽ©į, 4žčúÍįĄ ŽŹôžēą ŽįįžĖĎ ŪõĄ MUC4 mRNAžĚė ŽįúŪėĄ ž†ēŽŹĄŽ•ľ RTPCRŽ°ú žł°ž†ēŪēú Í≤įÍ≥ľ MUC4 mRNAžĚė ŽįúŪėĄžĚī ŽĆÄž°įÍĶįÍ≥ľ ŽĻĄÍĶźŪēėžėĞ̥ ŽēĆ Ž™®ŽĎź žĚėŽĮł žěąÍ≤Ć ÍįźžÜĆŪē®žĚĄ ŪôēžĚłŪēėžėÄŽč§(Fig. 2E).
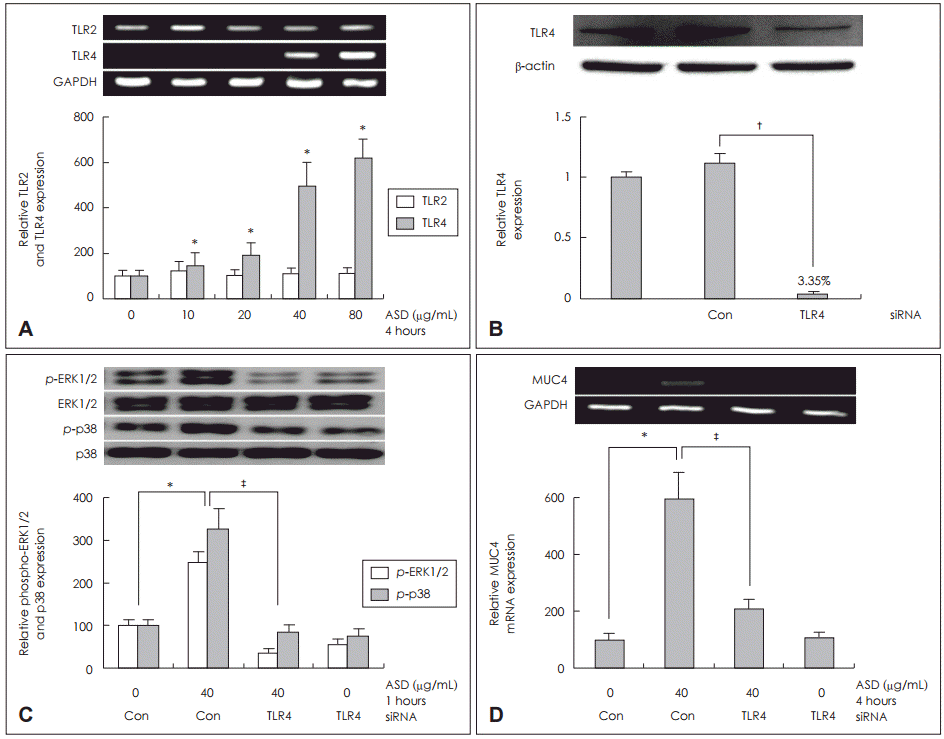
NCI-H292 žĄłŪŹ¨žóźžĄú Ūô©žā¨ÍįÄ TLR4Ž•ľ ŪÜĶŪēú MUC4 ŽįúŪėĄžóź ŽĮłžĻėŽäĒ žėĀŪĖ•
Ūô©žā¨žóź žĚėŪēī žú†ŽŹĄŽźú MUC4 ŽįúŪėĄžóź ŽĆÄŪēú TLRžĚė žó≠Ūē†žĚĄ ŪôēžĚłŪēėÍłį žúĄŪēī TLR2 mRNAžôÄ TLR4 mRNAžĚė ŽįúŪėĄžĚĄ RTPCRžĚĄ ŪÜĶŪēī Ž∂ĄžĄĚŪēėžėÄŽč§. Ūô©žā¨ÍįÄ TLR4 mRNAžĚė ŽįúŪėĄžĚĄ ž¶ĚÍįÄžčúžľįÍ≥†, žĚīŽäĒ žö©ŽüČžĚėž°īžĄĪžĚĄ Ž≥īžėÄŽč§. Í∑łŽü¨Žāė TLR2 mRNA ŽįúŪėĄžĚÄ ŽĆÄž°įÍĶįÍ≥ľ ŽĻĄÍĶźŪēėžėĞ̥ ŽēĆ žį®žĚīÍįÄ žóÜžóąŽč§(Fig. 3A).
Ūô©žā¨žóź žĚėŪēī žú†ŽŹĄŽźú MUC4 ŽįúŪėĄžóźžĄú TLR4žôÄ ERK1/2 MAPK, p38 MAPKžĚė žč†Ūėłž†ĄŽč¨ Í≤ĹŽ°úŽ•ľ ŪôēžĚłŪēėÍłį žúĄŪēėžó¨ TLR4 siRNAŽ•ľ žĚīžö©Ūēėžó¨ knockdownžčúŪā® ŪõĄ Western blotžúľŽ°ú Ž∂ĄžĄĚŪēėžėÄžúľŽ©į ž∂©Ž∂ĄŪēú Ūėēžßą ž†ĄŪôėžĚī žĚľžĖīŽā¨žĚƞ̥ ŪôēžĚłŪēėžėÄÍ≥†(Fig. 3B), Ūô©žā¨ 40 őľgžĚĄ Ūą¨žó¨ŪēėžėÄžúľŽ©į 1žčúÍįĄ ŽŹôžēą ŽįįžĖĎŪēú ŪõĄ žčúŪĖČŪēú Western blot Í≤Äžā¨ Í≤įÍ≥ľ ERK1/2 MAPKžôÄ p38 MAPKžĚė žĚłžāįŪôĒÍįÄ ŽĆÄž°įÍĶįÍ≥ľ ŽĻĄÍĶźŪēėžėĞ̥ ŽēĆ žĚėŽĮł žěąÍ≤Ć ÍįźžÜĆŪēėžėÄÍ≥†(Fig 3C), 4žčúÍįĄ ŽŹôžēą ŽįįžĖĎŪēú ŪõĄ MUC4 ŽįúŪėĄž†ēŽŹĄŽ•ľ RT-PCRŽ°ú Ž∂ĄžĄĚŪēú Í≤įÍ≥ľ MUC4 mRNA ŽįúŪėĄžĚī ŽĆÄž°įÍĶįÍ≥ľ ŽĻĄÍĶźŪēėžėĞ̥ ŽēĆ žĚėŽĮł žěąÍ≤Ć ÍįźžÜĆŪē®žĚĄ ŪôēžĚłŪēėžėÄŽč§(Fig. 3D).
ŪēėŽĻĄÍįĎÍįú ž†źŽßČ žÉĀŪĒľžĄłŪŹ¨žóźžĄú Ūô©žā¨ÍįÄ MUC4 ŽįúŪėĄžóź ŽĮłžĻėŽäĒ žėĀŪĖ•
žĚľžį® ŽįįžĖĎŽźú ŪēėŽĻĄÍįĎÍįú ž†źŽßČ žÉĀŪĒľžĄłŪŹ¨žóźžĄú Ūô©žā¨žĚė žö©ŽüČžóź ŽĒįŽ•ł TLR4 mRNAžôÄ MUC4 mRNA ŽįúŪėĄžóź ŽĮłžĻėŽäĒ žėĀŪĖ•žóź ŽĆÄŪēī žēĆžēĄŽ≥īÍ≥†žěź ŪēėŽĻĄÍįĎÍįú ž†źŽßČ žÉĀŪĒľžĄłŪŹ¨žóź Žč§žĖĎŪēú žö©ŽüČžĚė Ūô©žā¨(0, 10, 20, 40, 80 őľg)Ž•ľ ž≤ėŽ¶¨ŪēėžėÄžúľŽ©į, 4žčúÍįĄ ŽŹôžēą ŽįįžĖĎŪēú ŪõĄ žčúŪĖČŪēú RT-PCR Í≤Äžā¨žóźžĄú TLR4 mRNAžôÄ MUC4 mRNAžĚė ŽįúŪėĄžĚī Ž™®Žď† žö©ŽüȞ󟞥ú žĚėŽĮł žěąÍ≤Ć ž¶ĚÍįÄŪē®žĚĄ ŪôēžĚłŪē† žąė žěąžóąŽč§(Fig. 4A). ŽėźŪēú MUC4 ŽčĻŽč®ŽįĪ žÉĚžĄĪ ž†ēŽŹĄŽ•ľ ŪôēžĚłŪēėÍłį žúĄŪēī žčúŪĖČŪēú ELISA Í≤Äžā¨žóźžĄúŽŹĄ žĚėŽĮł žěąŽäĒ ŽčĻŽč®ŽįĪ žÉĚžĄĪ ž¶ĚÍįÄŽ•ľ ŪôēžĚłŪē† žąė žěąžóąŽč§(Fig. 4B). ERK1/2 MAPKžĚė žĖĶž†úž†úžĚł U0126(2 őľM)Í≥ľ p38žĚė žĖĶž†úž†úžĚł SB203580(20 őľM)žúľŽ°ú ž†Ą ž≤ėžĻė ŪõĄ Ūô©žā¨ 40 őľgŽ•ľ Ūą¨žó¨ŪēėžėĞ̥ ŽēĆŽäĒ MUC4 mRNA ŽįúŪėĄÍ≥ľ ŽčĻŽč®ŽįĪ žÉĚžĄĪžĚī žĚėŽĮł žěąÍ≤Ć ÍįźžÜĆŪēėžėÄŽč§(Fig. 4C and D).
Í≥† žįį
Ūô©žā¨ŽäĒ Žß§ŽÖĄ ŽīĄž≤†žĚīŽ©ī ÍĶ≠ŽāīŽ•ľ ŪŹ¨Ūē®Ūēú ŽŹôžēĄžčúžēĄ žßÄžó≠žóźžĄú ž£ľÍłįž†ĀžúľŽ°ú ŽįúžÉĚŪēėŽäĒ ÍłįžÉĀŪėĄžÉĀžĚīŽč§. Ūô©žā¨ ŪėĄžÉĀžóźžĄú žÉĀŽĆÄž†ĀžúľŽ°ú ŪĀ¨ÍłįÍįÄ ŪĀį žě֞쟎吏ĚÄ ž£ľŽ≥ÄŽ∂Ä ŽįŹ ŽįúžõźžßÄžóź Ž®łŽ¨ľÍ≤Ć ŽźėÍ≥†, ÍĶ≠Žāīžóź žú†žěÖŽźėŽäĒ Ūô©žā¨ žěÖžěźžĚė ŪĀ¨ÍłįŽäĒ ŽĆÄŽ∂ÄŽ∂Ą 10 őľm žĚīŪēėžĚė ŽĮłžĄłŽ®ľžßÄ(particulate matter 10)žĚīŽč§. ŽĮłžĄłŽ®ľžßÄŽäĒ ŪĚ°žěÖ žčú ŪēėŽ∂ÄÍłįÍīÄžßÄ ŽįŹ ŪŹźžĚė ÍįÄžä§-ÍĶźŪôėŽ∂ÄŽ∂ĄÍĻĆžßÄ žĻ®žį©Ūēėžó¨ Žč§žĖĎŪēú Íłįž†ĄžĚĄ ŪÜĶŪēī ŪėłŪĚ°Íłį žÜźžÉĀžĚĄ žú†ŽįúŪēėŽäĒ Í≤ÉžúľŽ°ú žēĆŽ†§ž†ł žěąŽč§[6]. ŪäĻŪěą žĶúÍ∑ľ žóįÍĶ¨žóźžĄú Ūô©žā¨ÍįÄ žā¨ŽěĆ ŪėłŪĚ°Íłį žÉĀŪĒľžĄłŪŹ¨žóźžĄú MUC5BžôÄ MUC8 ž†źžē°žú†ž†ĄžěźŽ•ľ Í≥ľŽįúŪėĄžčúŪā§Í≥†[7], žā¨ŽěĆ ŽĻĄžö© žÉĀŪĒľžĄłŪŹ¨žóźžĄúŽŹĄ MUC4žôÄ MUC5AC, MUC5B, MUC8žĚė ž†źžē°žú†ž†Ąžěź Í≥ľŽįúŪėĄžĚĄ žú†ŽįúŪēúŽč§Í≥† žēĆŽ†§ž†ł žěąŽč§[8]. Í∑łŽü¨Žāė, Ūô©žā¨ÍįÄ ŪėłŪĚ°Íłį Žį©žĖīÍłįž†ĄžĚė ž§ĎžöĒŪēú ž†źžÜƞ̳ MUC4žôÄ MUC16Í≥ľ ÍįôžĚÄ ŽßČÍ≤įŪē©Ūėē ž†źžÜĆžóź ŽĮłžĻėŽäĒ žėĀŪĖ•žóź ŽĆÄŪēīžĄúŽäĒ žóįÍĶ¨Žźú ŽįĒÍįÄ žóÜŽč§. žĚīžóź Ž≥ł žóįÍĶ¨žóźžĄúŽäĒ žā¨ŽěĆžĚė ŪėłŪĚ°Íłį žÉĀŪĒľžĄłŪŹ¨žóźžĄú Ūô©žā¨ÍįÄ ž£ľžöĒ ŽßČÍ≤įŪē©Ūėē ž†źžÜƞ̳ MUC4žôÄ MUC16žĚė ŽįúŪėĄžóź ŽĮłžĻėŽäĒ žėĀŪĖ•Í≥ľ ÍīÄŽ†® Íłįž†Ąžóź ŽĆÄŪēī ž§Ďž†źžĚĄ ŽĎźÍ≥† žóįÍĶ¨Ž•ľ žßĄŪĖČŪēėžėÄžúľŽ©į, Ž≥ł žóįÍĶ¨žĚė Í≤įÍ≥ľŽ•ľ ŪÜĶŪēī Ūô©žā¨ÍįÄ MUC4žĚė Í≥ľŽįúŪėĄžĚĄ žú†ŽįúŪē®žĚĄ ŪôēžĚłŪē† žąė žěąžóąŽč§.
ŪėłŪĚ°ÍłįžóźžĄú ž†ēžÉĀž†ĀžúľŽ°ú ž°īžě¨ŪēėŽäĒ MUC4ŽäĒ MUC1, MUC16Í≥ľ Ūē®ÍĽė ŪėłŪĚ°Íłį ž†źžē°žłĶžĚė ŽāīžłĶžĚĄ ÍĶ¨žĄĪŪēėŽäĒ ž£ľžöĒ ž†źžÜĆŽ°ú žôłŽ∂ÄŽ¨ľžßąÍ≥ľ žÜźžÉĀžúľŽ°úŽ∂ÄŪĄį ŪėłŪĚ°Íłį ž†źŽßČŽ•ľ Ž≥īŪėłŪēėŽäĒ ÍłįŽä•žĚĄ ŪēėŽäĒ ž§ĎžöĒŪēú ŽßČÍ≤įŪē©Ūėē ŽčĻŽč®ŽįĪžßąŽ°ú žēĆŽ†§ž†ł žěąŽč§[3]. ŽėźŪēú MUC4ŽäĒ žÉĀŪĒľžĄłŪŹ¨ žě¨žÉĀ(epithelial cell renewal), žĄłŪŹ¨ Ž∂ĄŪôĒ(differenciation), žĄłŪŹ¨ žč†Ūėłž†ĄŽč¨(cell signaling), žĄłŪŹ¨ Ž∂Äžį©(cell adhesion), žēĒžĚė ŽįúžÉĚ(cancer development) ŽďĪžóź ÍīÄžó¨ŪēėŽäĒ Í≤ÉžúľŽ°ú žēĆŽ†§ž†ł žěąŽč§. ŪäĻŪěą ŪėłŪĚ°Íłį žßąŪôėžóźžĄúŽäĒ žĄ¨žēĒžĘÖ(adenocarcinoma)žóźžĄú ŽÜížĚÄ ŽįúŪėĄžĚĄ Ž≥īžĚīŽ©į, žĄ¨žēĒžĘÖžĚė ÍįźŽ≥ĄžßĄŽč®Í≥ľ žėąŪõĄ ŪŹČÍįÄžóź žú†žö©Ūēú ŽįĒžĚīžė§ŽßąžĽ§Ž°ú žā¨žö©ŽźėÍ≥† žěąŽč§[11]. ŽėźŪēú, Žā≠žĄĪ žĄ¨žú†ž¶Ě(cystic fibrosis)Í≥ľ ŽßĆžĄĪŪŹźžáĄžĄĪŪŹźžßąŪôė(chronic obstructive pulmonary disease) ŽďĪžĚė ŽßĆžĄĪ žóľž¶ĚžĄĪ ŪėłŪĚ°Íłį žßąŪôėžóźžĄúŽŹĄ ž†ēžÉĀÍ≥ľ Žč§Ž•ł ŽįúŪėĄžĖĎžÉĀžĚĄ Ž≥īžĚīŽäĒ Í≤ÉžúľŽ°ú žēĆŽ†§ž†ł žěąŽč§[12,13]. Í∑łŽü¨Žāė žēĄžßĀÍĻĆžßÄ MUC4Ž•ľ ŪŹ¨Ūē®Ūēú Žč§žĖĎŪēú ŽßČÍ≤įŪē©Ūėē ž†źžÜĆžĚė žÉĚŽ¶¨ž†Ā ÍłįŽä•Í≥ľ Í≥ľŽįúŪėĄžĚī ÍįÄžßÄŽäĒ Ž≥ĎŪÉúžÉĚŽ¶¨Ūēôž†Ā žĚėŽĮłžóź ŽĆÄŪēī žĚīŪēīŪēėÍłį žúĄŪēīžĄúŽäĒ ž∂ĒÍįÄž†ĀžĚł žóįÍĶ¨ÍįÄ ŪēĄžöĒŪēú žč§ž†ēžĚīŽč§.
TLRžĚÄ ŪėłŪĚ°Íłį žÉĀŪĒľžĄłŪŹ¨žóźžĄú žôłŽ∂ÄŽ°úŽ∂ÄŪĄį žú†žěÖŽźėŽäĒ Ž≥Ďžõźž≤īŽ•ľ žĚłžčĚŪēėžó¨ žĄ†ž≤úŽ©īžó≠(innate immunity) ŽįėžĚĎžóź ž§ĎžöĒŪēú žó≠Ūē†žĚĄ ŽčīŽčĻŪēėŽ©į, ž†źžÜĆ Ž∂ĄŽĻĄ ž°įž†ąžóź žěąžĖīžĄúŽäĒ MUC4žôÄ MUC5B, MUC8 ŽįúŪėĄ ž¶ĚÍįÄžóź ž§ĎžöĒŪēú žó≠Ūē†žĚĄ ŪēúŽč§[7,9,10,14]. Ž≥ł žóįÍĶ¨žóźžĄúŽŹĄ žā¨ŽěĆ ŪėłŪĚ°Íłį žÉĀŪĒľžĄłŪŹ¨žóźžĄú Ūô©žā¨žóź žĚėŪēú TLR4 ŽįúŪėĄ žó¨Ž∂ÄŽ•ľ žēĆžēĄŽ≥ł Í≤įÍ≥ľ Ūô©žā¨ÍįÄ TLR4 ŽįúŪėĄžĚĄ žú†ŽŹĄŪēúŽč§ŽäĒ Í≤ɞ̥ žēĆ žąė žěąžóąŽč§.
žā¨ŽěĆžĚė ŪėłŪĚ°Íłį žÉĀŪĒľžĄłŪŹ¨žóźžĄú ž£ľžöĒ ž†źžē°žú†ž†Ąžěź ŽįúŪėĄžĚė žč†Ūėłž†ĄŽč¨ Í≤ĹŽ°ú ž§Ď ERK1/2 MAPKžôÄ p38 MAPKžóź ŽĆÄŪēú žóįÍĶ¨Žď§žĚĄ žāīŪéīŽ≥īŽ©ī, spleen tyrosine kinasežôÄ delphinidinžĚī ERK1/2 MAPK ŽėźŽäĒ p38 MAPK žč†Ūėłž†ĄŽč¨ Í≤ĹŽ°úŽ•ľ ŪÜĶŪēī MUC5AC ŽįúŪėĄÍ≥ľ MUC5B ŽįúŪėĄ, MUC8 ŽįúŪėĄžĚĄ žú†ŽŹĄŪēėžėÄžúľŽ©į[14,15], ő≤-glucanÍ≥ľ phorbol 12-myristate 13-acetateŽäĒ p38 MAPK žč†Ūėłž†ĄŽč¨ Í≤ĹŽ°úŽ•ľ ŪÜĶŪēī MUC4 ŽįúŪėĄžĚĄ žú†ŽŹĄŪēėžėÄÍ≥†[10,16], ŪäĻŪěą Ūô©žā¨ÍįÄ ERK2 MAPKžôÄ p38 MAPK žč†Ūėłž†ĄŽč¨ Í≤ĹŽ°úŽ•ľ ŪÜĶŪēī MUC5BžôÄ MUC8 ŽįúŪėĄžĚĄ žú†ŽŹĄŪēėžėÄŽč§[7]. žĚīžóź Ž≥ł žóįÍĶ¨žóźžĄúŽŹĄ Ūô©žā¨ÍįÄ MUC4 ŽįúŪėĄ žú†ŽŹĄžóź žěąžĖī ERK1/2 MAPKžôÄ p38 MAPK žč†Ūėłž†ĄŽč¨ Í≤ĹŽ°úÍįÄ ž§ĎžöĒŪēú žó≠Ūē†žĚĄ Ūē† Í≤ÉžúľŽ°ú žÉĚÍįĀŽźėžĖī ŪôēžĚłŪēī Ž≥ł Í≤įÍ≥ľ Ūô©žā¨ÍįÄ ERK1/2 MAPKžôÄ p38 MAPK žĚłžāįŪôĒŽ•ľ žĚėŽĮł žěąÍ≤Ć ž¶ĚÍįÄžčúžľįÍ≥†, ERK1/2 MAPKžôÄ p38 MAPK žĚłžāįŪôĒŽ•ľ žĖĶž†úŪēėžėĞ̥ Í≤Ĺžöį Ūô©žā¨žóź žĚėŪēú MUC4 ŽįúŪėĄžĚī ÍįźžÜĆŽź®žĚĄ ŪôēžĚłŪē† žąė žěąžóąŽč§. žĚīŽ•ľ ŪÜĶŪēī Ūô©žā¨ÍįÄ ERK1/2 MAPKžôÄ p38 MAPK žč†Ūėłž†ĄŽč¨ Í≤ĹŽ°úŽ•ľ ŪÜĶŪēī MUC4 ŽįúŪėĄžĚĄ ž¶ĚÍįÄžčúŪā§ŽäĒ Í≤ɞ̥ žēĆ žąė žěąžóąŽč§. ŽėźŪēú Ūô©žā¨žóź žĚėŪēú MUC4 ŽįúŪėĄžĚė ž¶ĚÍįÄžóź žěąžĖīžĄú TLR4, ERK1/2 MAPK, p38 MAPKžĚė žóįÍīÄžĄĪžĚĄ ŪôēžĚłŪēī Ž≥ł Í≤įÍ≥ľ Íłįž°īžĚė žóįÍĶ¨Žď§Í≥ľ žú†žā¨ŪēėÍ≤Ć TLR4žóź Žß§ÍįúŽźú ERK1/2 MAPKžôÄ p38 MAPK žč†Ūėłž†ĄŽč¨ ž≤īÍ≥ĄžĚė ŪôúžĄĪŪôĒÍįÄ žóįÍīÄŽźėžĖī žěąžĚƞ̥ ŪôēžĚłŪē† žąė žěąžóąŽč§(Fig. 5). Í∑łŽü¨Žāė Ž≥ł žóįÍĶ¨žĚė ž†úŪēúž†źžĚÄ ŽßČÍ≤įŪē©žĄĪ ž†źžÜĆžĚė Ž∂ĄžĄĚžóźžĄú MUC1žĚė ŽįúŪėĄŽ≥ÄŪôĒŽ•ľ ŪôēžĚłŪēėžßÄ Ž™ĽŪēėžėĎ觎äĒ ž†ź, Ž≥ĶŪē©Ž¨ľžĚł Ūô©žā¨žĚė žĄĪŽ∂Ąžóź ŽĆÄŪēú ÍįúŽ≥Ąž†Ā Ūö®Í≥ľžóź ŽĆÄŪēú Í≤Äž¶ĚžĚī Ž∂Äž°ĪŪēėŽč§ŽäĒ ž†źÍ≥ľ žĄłŪŹ¨žóźŽßĆ ÍĶ≠ŪēúŽźú žč§Ūóė Í≤įÍ≥ľžĚīŽĮÄŽ°ú ž∂ĒÍįÄž†ĀžĚł ŽŹôŽ¨ľžč§ŪóėžĚĄ ŪÜĶŪēī žóįÍĶ¨ Í≤įÍ≥ľžóź ŽĆÄŪēú ŪôēžĚłžĚī ŪēĄžöĒŪēėŽč§ŽäĒ ž†źžĚĄ Žď§ žąė žěąÍ≤†Žč§. žĚīŽ•ľ ŪēīÍ≤įŪēėÍłį žúĄŪēīžĄú ž∂ĒÍįÄž†ĀžĚł žóįÍĶ¨ÍįÄ Ží§ŽĒįŽĚľžēľ Ūē† Í≤ÉžúľŽ°ú žÉĚÍįĀŽźúŽč§.
Í≤įŽ°†ž†ĀžúľŽ°ú Ž≥ł žóįÍĶ¨ Í≤įÍ≥ľŽ•ľ ŪÜĶŪēī Ūô©žā¨ÍįÄ TLR4žóź žĚėž°īŪēú ERK1/2 MAPKžôÄ p38 MAPK žč†Ūėłž†ĄŽč¨ Í≤ĹŽ°úŽ•ľ ŪÜĶŪēī MUC4 ŽįúŪėĄžĚĄ ž¶ĚÍįÄžčúŪā§ŽäĒ Í≤ɞ̥ žēĆ žąė žěąžóąÍ≥†, žĚīŽäĒ Ūô©žā¨žôÄ ÍīÄŽ†®Žźú Žč§žĖĎŪēú ŪėłŪĚ°Íłį žßąŪôėžĚė Ž≥ĎŪÉúžÉĚŽ¶¨Ž•ľ žĚīŪēīŪēėŽäĒ Žćį ž§ĎžöĒŪēú ÍłįžīąžěźŽ£ĆÍįÄ Žź† Í≤ÉžúľŽ°ú žÉĚÍįĀŽźúŽč§.